 夜雨聆风
夜雨聆风在植物体内,接续韧皮部装载的是一场规模宏大、精密无比、日夜不息地运转着的“跨省快递”。它的起点是叶片——光合作用的“制造工厂”;终点可能是几十厘米外的嫩芽、几米高处的果实,或是深埋地下的块茎块根,甚至新根和根毛。而承运这条生命专线的,正是由活细胞构成的韧皮部。我们已经知道,糖分要进入这条专线,需先完成一次精妙的“装车”(韧皮部装载,见前文)。但同样具有挑战性的是,这些糖如何完成一场“跨省”奔袭?抵达目的地后,又如何被安全、高效、甚至“按需分配”地卸下?这背后,藏着植物对资源调度最深刻的生存智慧。
这场长途运输的驱动力,出人意料地朴素——它靠的不是生物引擎,而是一个高效的物理引擎:压力差。想象你在软管一端加压,水自然流向低压端。植物正是如此操作。如下图所示,在叶片(源),糖被主动泵入叶脉韧皮部筛分子,导致内部溶质浓度飙升、水势骤降。于是,邻近木质部导管内的水分迅速渗入,使筛分子原生质体膨胀增压。而在根、果实或嫩叶(库),糖被快速取走利用或储存,筛分子内浓度下降,水势升高,水分流出,压力随之降低。这一高一低之间,形成稳定的压力梯度,推动满载蔗糖等分子的汁液,在韧皮部筛分子中以每小时约一米的速度,从源直抵库。
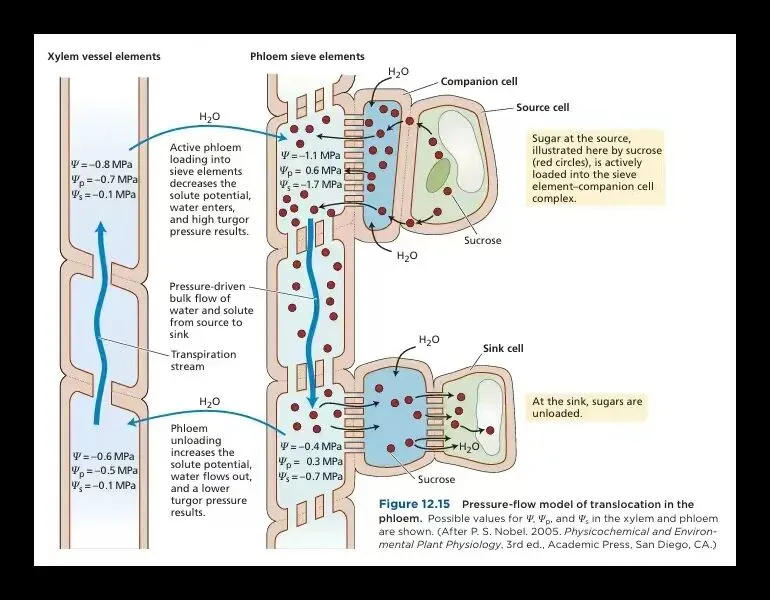
有人会问:如果库在上方(比如顶芽),而木质部的水主要靠蒸腾向上拉,那此时木质部的水怎么“逆水势而下”完成模型所需的“水分循环”?这正是压力流动模型本身的小“瑕疵”,尤其在蒸腾微弱区域(嫩芽或果实附近)的局部水势动态仍存在理论局限。该模型能很好地描述源在上库在下的长距离运输的宏观驱动力,但对卸载端位于上方时的复杂的细胞间水分再分配,尚需结合其他机制补充完善。
如果说长途运输是一场依靠物理法则的“被动漂流”,那么卸货则是一场高度智能化的“精准投递”。植物绝不会用同一种方式处理所有订单。面对正在分裂生长的嫩芽和根尖或根毛这类如饥似渴的“急单客户”,系统启动共质体卸载模式。如下图(A)所示,糖分通过细胞间的胞间连丝,像水流过连通水管一样,直接扩散到周围细胞。这是一种顺浓度梯度的被动卸载过程,响应快、效率高。
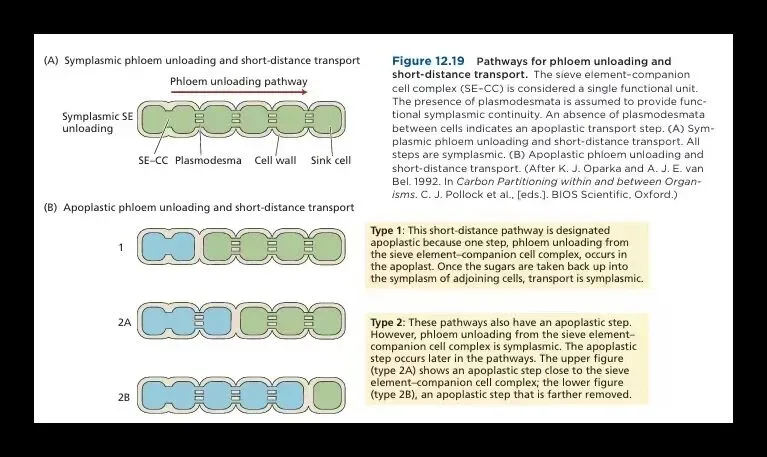
但当目的地是甜菜的块根、甘蔗的茎秆、灌浆中的种子或正在长大的果实时,订单性质就变了——客户要的不是“吃掉”,而是“囤积”。此时,系统切换至质外体卸载模式。如上图(B)所示,糖先被释放到细胞壁间隙(相当于“公共分拣站”),再由库细胞主动“摄入”。
这一库细胞的主动“摄入”,有两种截然不同的策略,分别适配不同库器官的代谢需求。
第一种是“水解–单糖入库”模式,常见于桃、樱桃、番茄等肉质果实。蔗糖从筛分子-伴胞复合体卸入质外体后,立即被细胞壁酸性转化酶水解为葡萄糖和果糖。这些单糖随后由 MST/STP 家族转运蛋白(H⁺-己糖同向转运体)主动转运进入库细胞——该过程依赖质膜 H⁺-ATPase 建立的质子梯度,属于次级主动运输。
第二种是“蔗糖直送–双糖入库”模式,典型见于大豆种子、甜菜块根等储藏器官。此处质外体中几乎无转化酶活性,蔗糖以完整形式存在于细胞间隙。库细胞则通过 SUT/SUC 家族转运蛋白(H⁺-蔗糖同向转运体),利用同样的质子驱动力,将蔗糖逆浓度梯度泵入胞内,直接用于合成淀粉或储存。此路径无需水解步骤,能量效率高,适合大规模囤积。
两种模式在卸载与入库环节高度协同、互不交叉:水解路径配单糖转运,保留路径配蔗糖转运。正是这种分子层面的精准匹配,使植物能按需调度碳流,实现从“即时消费”到“战略储备”的灵活切换。
从源端的压力驱动长距离运输,到库端依需求分化的卸载策略,再到由特异转运蛋白执行的精准入库,植物构建了一套高效、可塑且高度协调的碳分配系统。这套系统既依赖物理规律的普适性,又通过分子机器的多样性实现精细调控,最终将光合产物精准送达生长、储存或繁殖所需之处——这不仅是物质的流动,更是生命对资源最优配置的无声智慧(详细配送和分享机制下文详解)。
