 夜雨聆风
夜雨聆风





NBT | 新型DNA碱基编辑器–基于ADAR脱氨酶的单核苷酸分辨率DNA腺嘌呤碱基snuABE
传统腺嘌呤碱基编辑器虽应用广泛且编辑效率高,但旁观者编辑问题严重制约其治疗应用。2026年7月10日,韩国成均馆大学Sangsu Bae团队在Nature Biotechnology上发表题为“Engineered ADARs enable precision A-to-G base editing of DNA”的研究论文。该研究开发了单核苷酸分辨率腺嘌呤碱基编辑器snuABE,将ADAR脱氨酶结构域与nCas9-H840A融合,通过引入靶标特异性错配的tagRNA,实现了对DNA:RNA杂合双链中目标腺嘌呤的高特异性A-to-G编辑。通过跨物种ADAR筛选、La蛋白3’端保护、AI驱动定向进化(EvolvePro)及额外切口和MLH1-SB介导的错配修复抑制等系统性优化,snuABE4.2n在HEK293T细胞中实现中位5.4%、最高50.0%的编辑效率,且无DNA脱靶编辑。在VHL综合征模型中实现高达63.6%的无旁观者精准校正,并在小鼠肝脏中验证了体内适用性。

主要研究结果
1,通过将ADAR脱氨酶结构域与nCas9-H840A融合,并设计引入错配的tagRNA,实现DNA:RNA杂合双链中靶标腺嘌呤的单核苷酸分辨率A-to-G编辑。
传统ABEs使用TadA脱氨酶作用于单链DNA,而snuABE利用ADAR(天然作用于双链RNA的腺苷脱氨酶)在DNA:RNA杂合双链中的活性。研究者将人ADAR2dd(含E488Q高活性突变)融合至nCas9-H840A的N端,并设计3’末端延伸的tagRNA以在靶标腺嘌呤处引入特定错配(图1a-b)。在HEK293T细胞FANCF位点测试显示,ABE8e编辑多个腺嘌呤(A12、A13、A18效率达61.0%),而snuABE1.0仅选择性编辑A12(1.1%)(图1c)。为提升活性,从20个物种中筛选22种ADARdd直系同源物,人虱(Pediculus humanus)来源PhADARdd在AAVS1位点达6.0%编辑,命名为snuABE2.0(图1d及补充图1)。
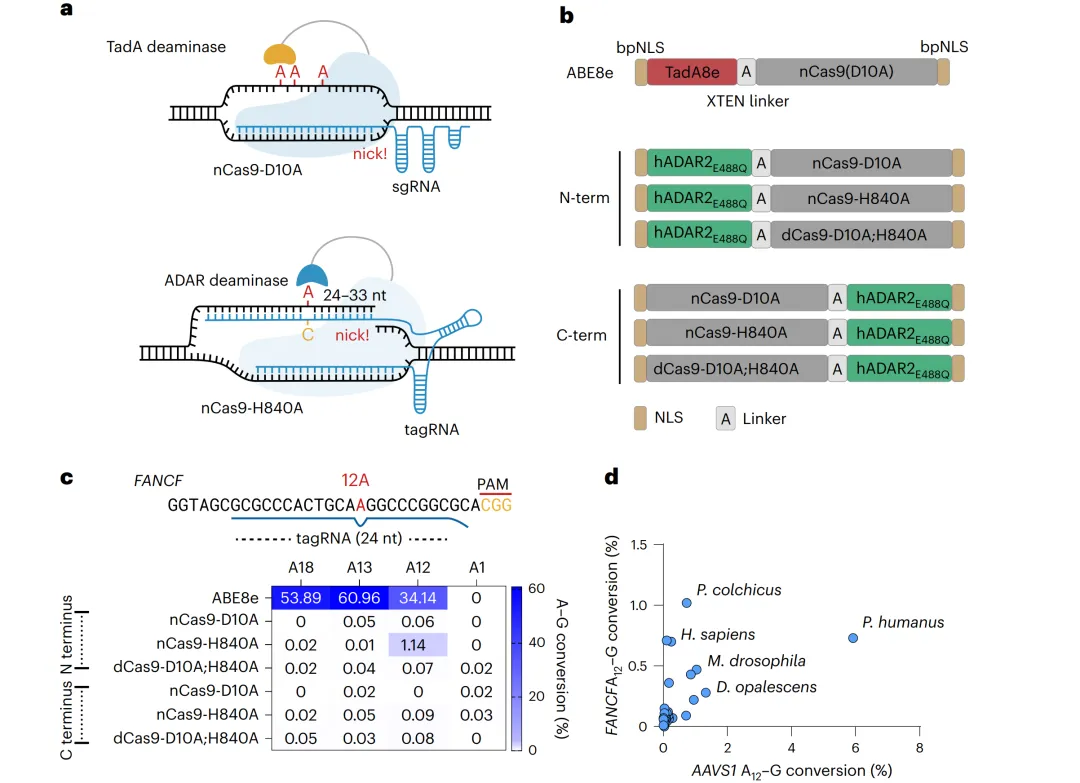
图1 snuABE的概念设计与跨物种ADAR筛选
2,通过PEmax启发工程化、La蛋白3’端保护及tagRNA长度优化,snuABE3.1在AAVS1和FANCF位点分别达20.0%和12.1%编辑效率。
将PEmax中SV40内部NLS、nCas9-H840A的R221K/N394K突变和C端c-Myc NLS引入snuABE2.0,生成snuABE2.1(图2a)。考虑到tagRNA与pegRNA结构相似性,共表达La蛋白(全长或1-194截短体)保护tagRNA 3’端,AAVS1编辑效率升至8.6%(图2b-c)。将La(1-194)直接融合至snuABE2.0 C端,生成snuABE3.0,编辑效率进一步提升至16.5%;再引入snuABE2.1的NLS和突变,最终snuABE3.1在AAVS1和FANCF分别达20.0%和12.1%(图2b-c)。在DNMT1位点,针对第13或15位腺嘌呤设计特异性tagRNA,snuABE3.1分别实现5.3%和6.5%的精准单一位点编辑,旁观者编辑极低(<1.1%),而所有传统ABEs均编辑多个腺嘌呤(图2d)。tagRNA长度优化显示24–33 nt最佳(图3a及扩展数据图1),编辑窗口集中于12–16位(图3d及扩展数据图2),且偏好5’ A/T- C/G-3’(图3e)。
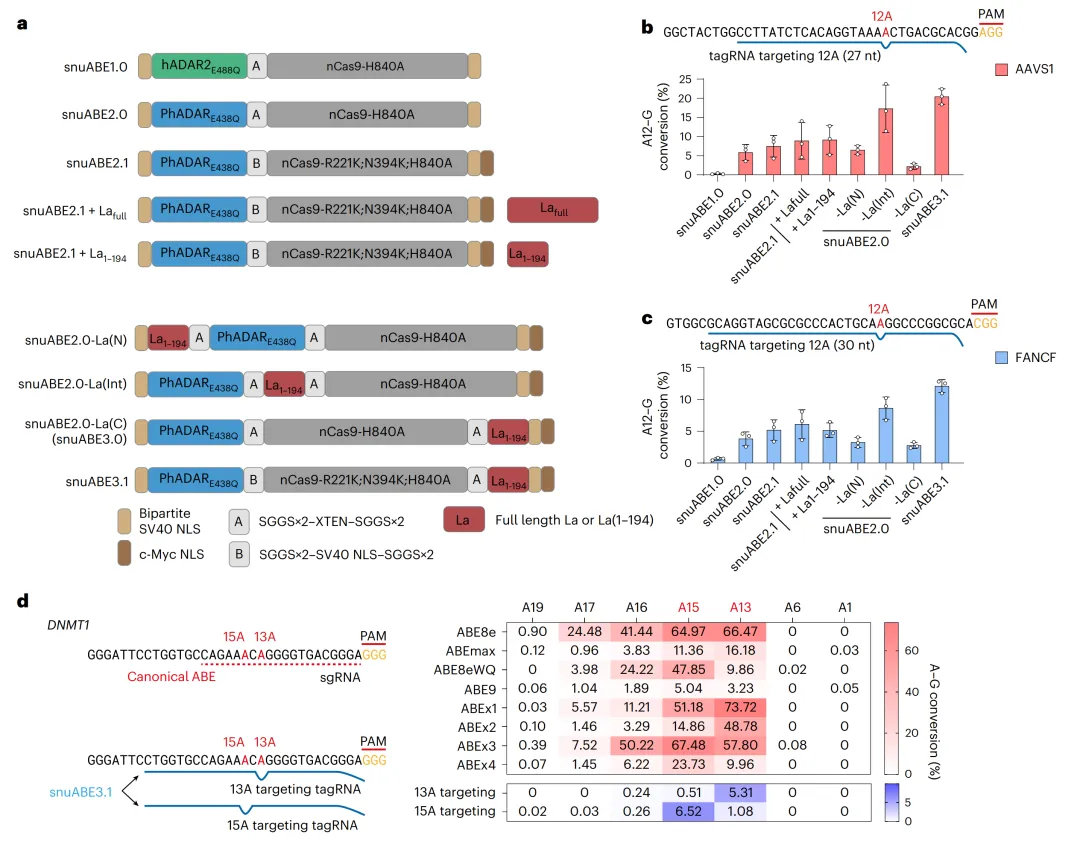
图2 snuABEs的工程化与tagRNA切换验证的单核苷酸选择性
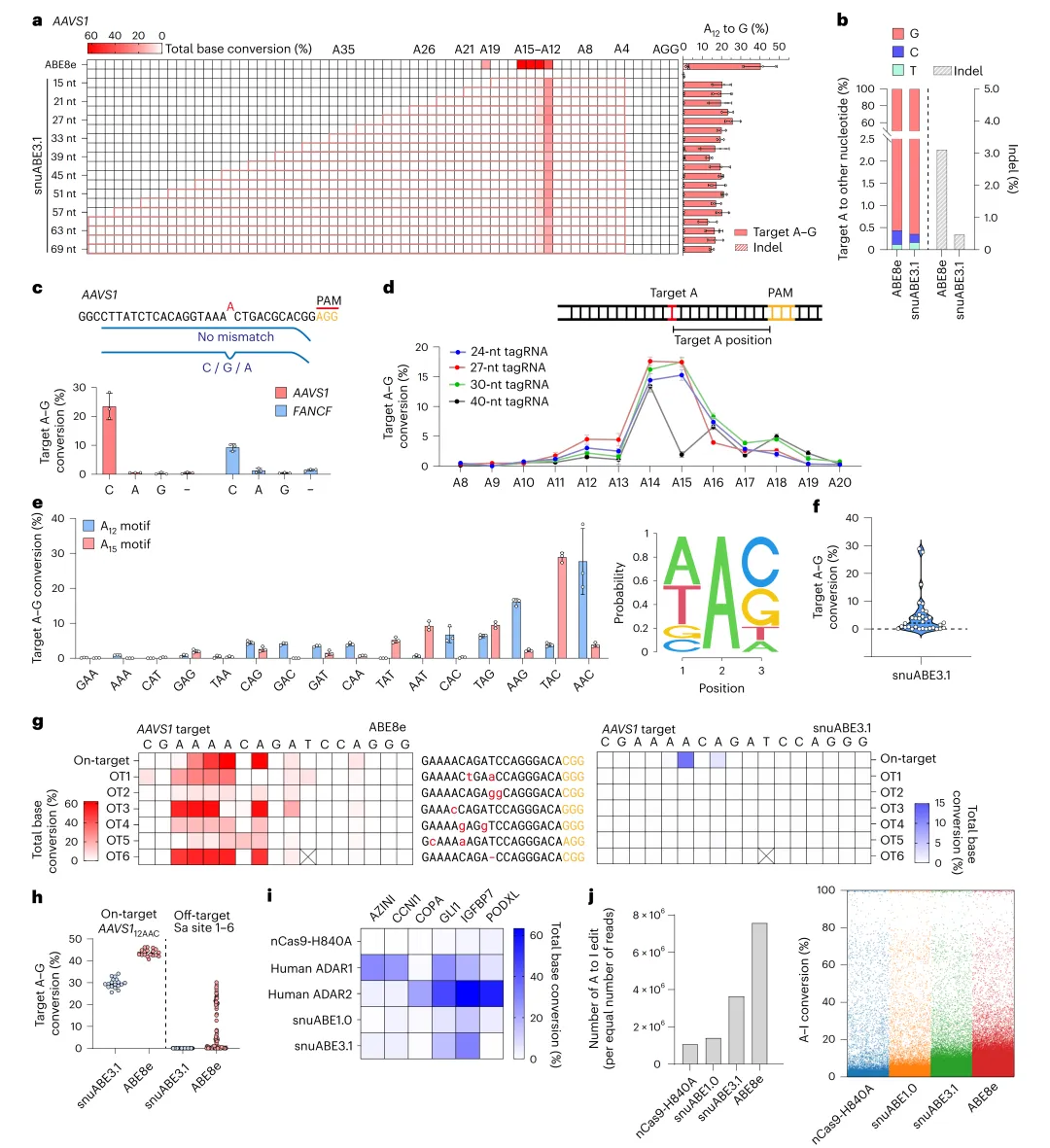
图3 snuABE3.1的功能表征
3,AI驱动EvolvePro定向进化结合额外切口和MLH1-SB介导的MMR抑制,snuABE4.2n中位编辑效率达5.4%、最高50.0%。
AlphaFold3预测PhADARdd结构显示N端flap可能非催化必需,截短10个残基后生成snuABE4.0,编辑效率提升(图4a-b)。采用EvolvePro进行五轮AI引导定向进化(图4c):首轮在5个内源靶点测试187个单点突变,每靶点选Top 8用于迭代训练;末轮预测Top 20多突变组合,其中含E438Q、I536S、E637F的snuABE4.1在多种序列背景下编辑效率显著提升,中位编辑率从2.1%(snuABE3.1)升至4.8%(图4d-g)。AlphaFold3显示I536S和E637F位于DNA:RNA结合环附近而非催化核心(扩展数据图6)。借鉴PE3策略引入对侧链额外切口(ngRNA),snuABE4.1n中位编辑率升至5.4%(图4g及扩展数据图7a-b),但ngRNA位置影响indel率(扩展数据图7c-d)。由于nCas9-H840A在切口链引入错配可能触发MMR,共表达MLH1dn或MLH1-SB(小分子结合剂)抑制MMR,在MMR正常细胞(HEK293、HeLa、U2OS)中编辑效率显著提升,而在MMR缺陷HEK293T中无改善(图4h及补充图2)。综合优化获得snuABE4.2n。DNA脱靶评估(6个Cas-OFFinder预测位点和正交R-loop assay)显示snuABE4.1无DNA脱靶编辑(图4i-j及扩展数据图5),但RNA脱靶随编辑活性提升而增加(图4k-l及补充图3-4)。在患者来源成纤维细胞和iPS细胞中分别达16.3%和1.4%编辑效率(图4m-n),PAM-flexible SpNG和SpRY变体亦有活性(补充图5)。
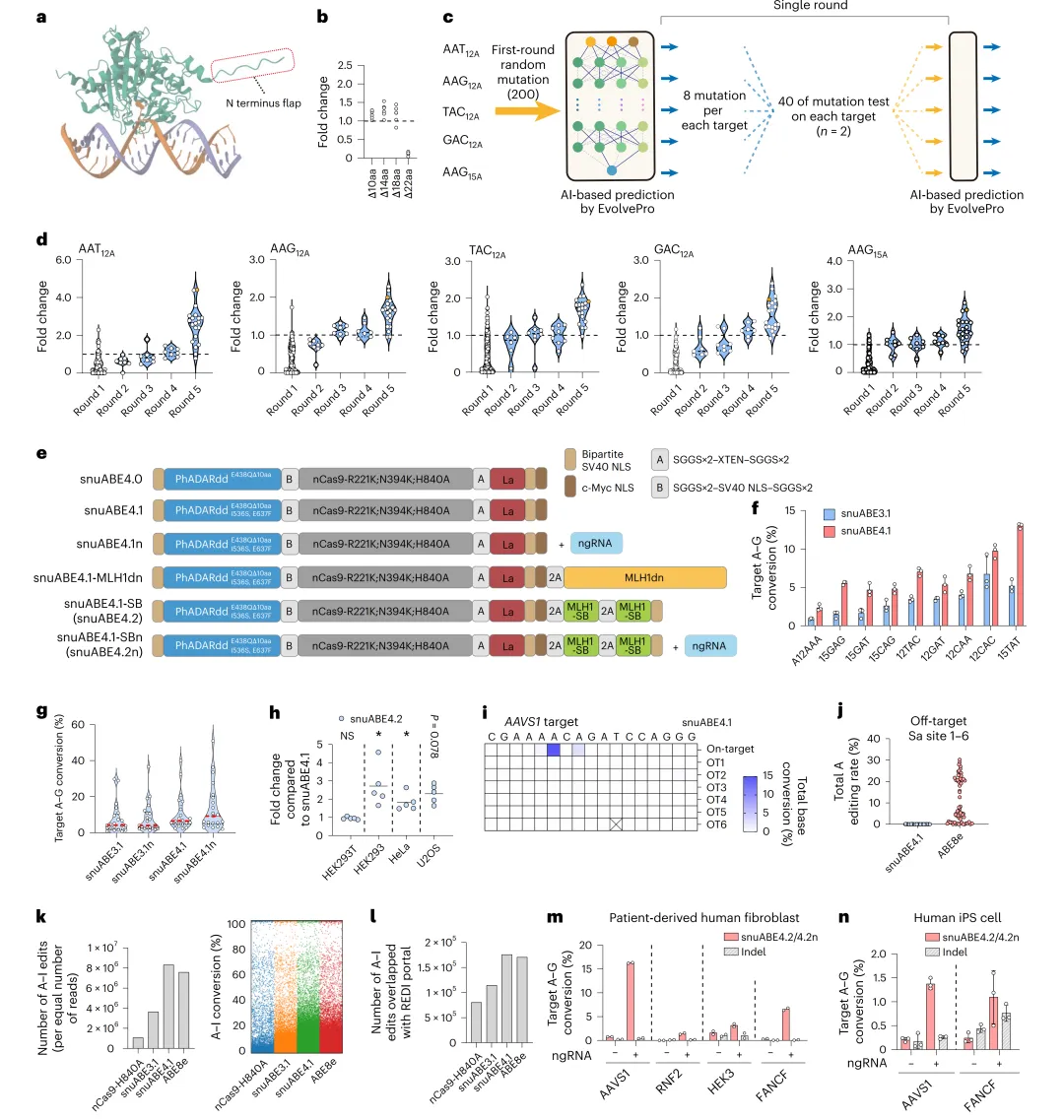
图4 AI引导进化与snuABE变体的工程化优化
4,snuABE在VHL综合征模型、PCSK9和BCL11A增强子中实现无旁观者精准编辑,并在小鼠肝脏中验证体内适用性。
在VHL综合征HEK293T细胞模型(c.332G>A)中,snuABE4.1实现高达63.6%的A-to-G校正且无旁观者编辑,而传统ABEs编辑窗口内多个腺嘌呤被修改(图5a-b)。在PCSK9外显子1和4剪接供体位点,snuABE4.2n分别达10.5%和12.3%编辑效率(图5c-d)。在BCL11A +55红系增强子(TGN-wGATAR基序),snuABE4.2n在K562细胞中达18.1%编辑效率且indel极低(图5e-f)。在NIH/3T3细胞中靶向小鼠Gcnt4内含子,snuABE4.1效率仅0.9%,而snuABE4.2n升至25.7%(补充图5)。通过水动力尾静脉注射将snuABE4.2n三质粒系统递送至小鼠肝脏,7天后平均编辑效率达2.0%(n=3),无旁观者编辑(图5g-i)。
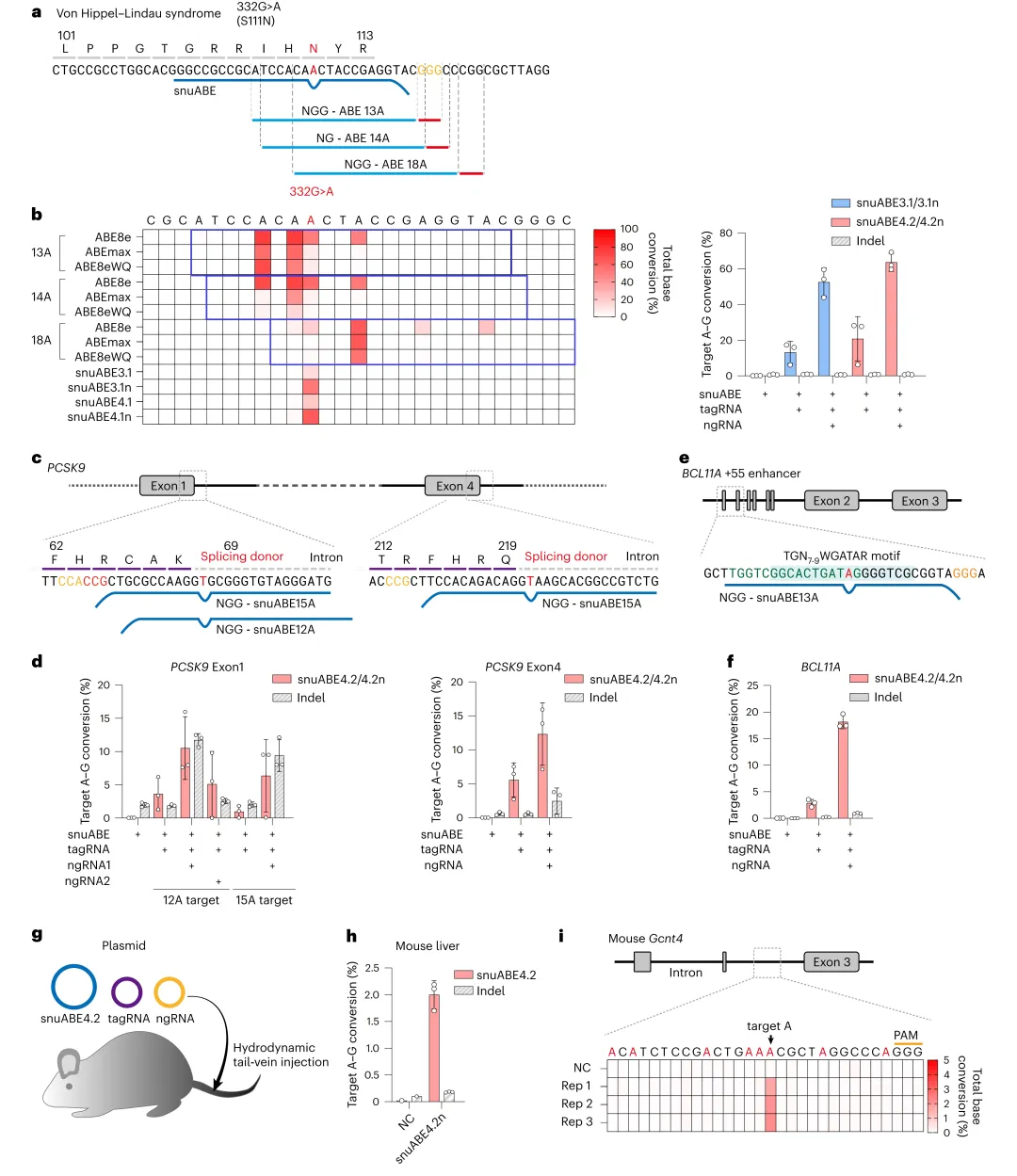
图5 snuABE在疾病模型和体内的应用
综上所述,本研究开发了snuABE系统,通过将ADAR脱氨酶结构域与nCas9-H840A融合并设计靶标特异性错配tagRNA,实现了DNA:RNA杂合双链中腺嘌呤的单核苷酸分辨率A-to-G编辑。通过跨物种ADAR筛选、La蛋白3’端保护、AI驱动EvolvePro定向进化及额外切口和MLH1-SB介导的MMR抑制等系统性优化,snuABE4.2n在中位编辑效率5.4%、最高50.0%的同时无DNA脱靶编辑,在VHL综合征模型中实现63.6%的无旁观者精准校正,并在小鼠肝脏中验证了体内适用性。然而,snuABE4.2n编辑效率仍低于传统ABE8e,且存在序列偏好性;AI进化引入的I536S和E637F突变在提升DNA编辑的同时增加了RNA脱靶编辑;tagRNA设计需针对每位点优化3’延伸长度(24–33 nt);目前主要依赖质粒共转染,递送系统有待进一步开发。
参考来源:
Im, H.W., Jeong, B., Lee, Y. et al. Engineered ADARs enable precision A-to-G base editing of DNA. Nat Biotechnol (2026). https://doi.org/10.1038/s41587-026-03223-z
免责声明:本文旨在传递基因编辑与治疗最新讯息,不代表平台立场,不构成任何投资意见和建议,以官方/公司公告为准。本文也不是治疗方案推荐,如需获得治疗方案指导,请前往正规医院就诊