一区 if:12.8
结果:
NeuroD1在胰腺癌中作为保护因子的作用
基因集GOBP上皮细胞分化的负调控包含一系列对上皮细胞分化发挥负向调控作用的基因,这些基因通过抑制或减缓分化过程来维持细胞的未分化状态或促进替代性细胞命运决定。本研究首先利用TCGA数据库,基于GOBP上皮细胞分化的负调控基因集计算每个TCGA胰腺癌转录组测序样本的单样本评分,从而预测每个样本的分化状态,并将其分为相对高分化组和相对低分化组。对患者预后数据的分析显示,相对高分化组的总生存期、疾病特异性生存期和无进展间隔期均显著优于相对低分化组。通过将GOBP上皮细胞分化的负调控基因集、预后差异基因以及编码特定亚细胞定位蛋白的基因进行交集分析,识别出54个可能参与上皮细胞分化和恶性行为的关键基因。随后整合并分析了公开可用的胰腺癌单细胞数据,依据先前的导管细胞注释方法,将导管细胞分为I型导管细胞和II型恶性导管上皮细胞。分析这54个关键基因在导管细胞中的表达水平与单样本分化评分之间的相关性,结果显示出显著关联。小提琴图展示了与单样本分化评分显著相关的前10个基因在I型正常导管细胞和II型恶性导管上皮细胞中的差异表达,观察到NeuroD1表达降低在II型恶性导管上皮细胞中最为显著。进一步的风险比分析表明NeuroD1在胰腺癌中作为保护因子。在恶性导管细胞中,NeuroD1表达与单样本分化评分呈负相关。对公共数据库数据的分析初步表明,NeuroD1高表达与胰腺癌的高分化表型相关,NeuroD1可能作为胰腺癌的保护因子发挥作用。
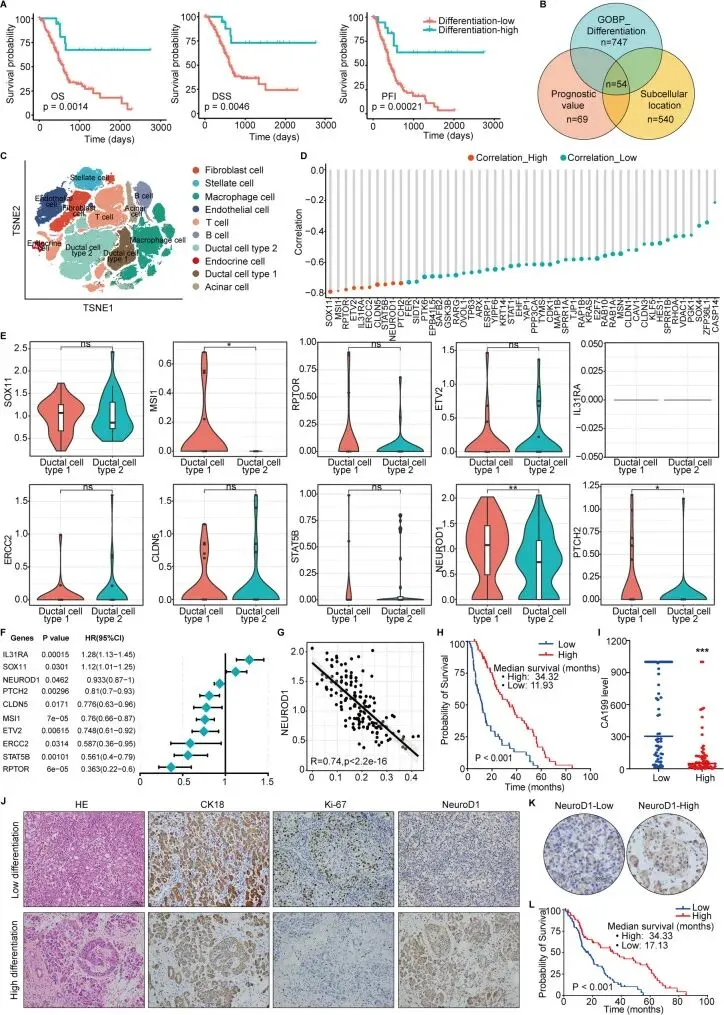 接着,对本中心治疗的60例高分化胰腺导管腺癌患者和60例低分化胰腺导管腺癌患者进行了生存分析。结果显示,高分化胰腺癌患者的生存时间显著长于低分化胰腺癌患者。此外,高分化组术前CA199水平显著较低。这些结果表明高分化胰腺癌确实表现出较低的恶性程度,并与较好的患者预后相关。为进一步探讨这一现象,使用IHC染色检测了CK18、Ki-67和NeuroD1在高分化和低分化胰腺导管腺癌中的表达水平,结果显示高分化胰腺癌表现出细胞增殖减少和NeuroD1蛋白表达增加,提示NeuroD1表达升高与胰腺癌细胞分化程度增加相关。随后量化了本中心另外120例胰腺导管腺癌患者中NeuroD1蛋白的表达,将其分为NeuroD1高表达组和低表达组。分析NeuroD1表达与患者生存时间的相关性发现,NeuroD1高表达患者的生存时间显著更长。结合公共数据库生物信息学分析结果及对患者组织样本的分析,可以得出结论:在胰腺癌患者中,NeuroD1表达与细胞分化水平升高及患者预后改善相关。这些发现提示NeuroD1可能作为胰腺癌的保护因子。
接着,对本中心治疗的60例高分化胰腺导管腺癌患者和60例低分化胰腺导管腺癌患者进行了生存分析。结果显示,高分化胰腺癌患者的生存时间显著长于低分化胰腺癌患者。此外,高分化组术前CA199水平显著较低。这些结果表明高分化胰腺癌确实表现出较低的恶性程度,并与较好的患者预后相关。为进一步探讨这一现象,使用IHC染色检测了CK18、Ki-67和NeuroD1在高分化和低分化胰腺导管腺癌中的表达水平,结果显示高分化胰腺癌表现出细胞增殖减少和NeuroD1蛋白表达增加,提示NeuroD1表达升高与胰腺癌细胞分化程度增加相关。随后量化了本中心另外120例胰腺导管腺癌患者中NeuroD1蛋白的表达,将其分为NeuroD1高表达组和低表达组。分析NeuroD1表达与患者生存时间的相关性发现,NeuroD1高表达患者的生存时间显著更长。结合公共数据库生物信息学分析结果及对患者组织样本的分析,可以得出结论:在胰腺癌患者中,NeuroD1表达与细胞分化水平升高及患者预后改善相关。这些发现提示NeuroD1可能作为胰腺癌的保护因子。NeuroD1抑制胰腺癌细胞的增殖
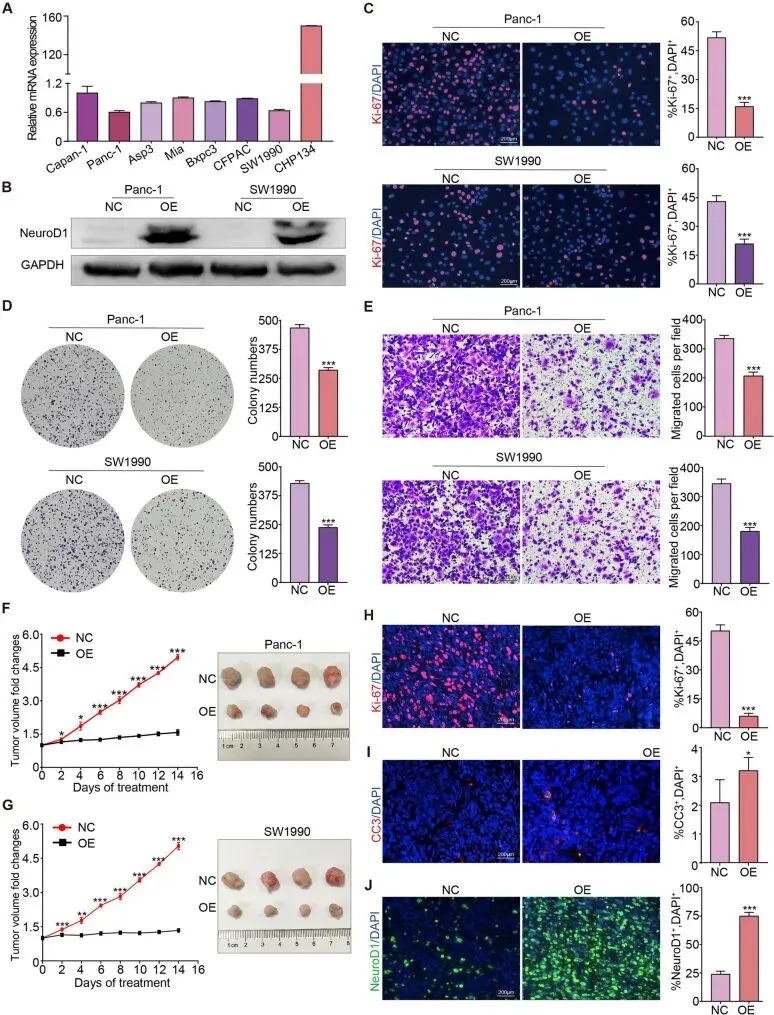
接下来探讨了NeuroD1在胰腺癌中的具体作用。通过RT-qPCR评估了NeuroD1在常见胰腺癌细胞系及人神经母细胞瘤细胞系CHP134中的表达水平。在胰腺癌细胞系中,Panc-1和SW1990细胞表现出最低的NeuroD1表达水平,因此选用这些细胞系通过慢病毒感染建立稳定的NeuroD1过表达细胞系,以空质粒转染的相同细胞系作为阴性对照。Western Blotting证实NeuroD1过表达细胞系已成功建立。
通过对增殖细胞标志物Ki-67蛋白进行免疫荧光染色以及集落形成实验,观察到NeuroD1过表达显著降低了胰腺癌细胞的增殖能力。Transwell实验显示NeuroD1还降低了胰腺癌细胞的迁移能力。通过将Panc-1和SW1990细胞接种裸鼠建立皮下肿瘤模型,发现NeuroD1过表达在体内阻碍了胰腺癌的进展。收集皮下肿瘤组织并进行Ki-67和Cleaved Caspase-3蛋白的免疫荧光染色,结果显示胰腺癌细胞的增殖潜能显著降低而凋亡增加。NeuroD1蛋白的免疫荧光染色证实了NeuroD1过表达模型的成功建立。综上所述,这些结果表明上调NeuroD1表达在体外和体内均能抑制胰腺癌细胞的增殖。NeuroD1诱导胰腺癌细胞转分化为神经元
NeuroD1过表达的胰腺癌细胞出现形态学改变,呈现出神经元样细胞形态。为评估NeuroD1过表达诱导的细胞变化,在诱导NeuroD1过表达后的第0、3和7天,使用CD56和CD326特异性抗体进行流式细胞术检测。结果显示,随着NeuroD1过表达的增加,CD326阳性细胞的百分比持续下降,而CD56阳性细胞的百分比持续上升,同时共表达CD326和CD56的细胞比例也增加,提示癌细胞正在逐步转分化为神经元。为探索这一转化的机制,对阴性对照Panc-1细胞和NeuroD1过表达Panc-1细胞进行了RNA测序分析。GSEA富集分析显示,在NeuroD1过表达细胞中,与DNA复制和线粒体功能相关的GSEA通路显著下调,表明促进细胞增殖和生长的生物学过程受到抑制;相反,TNF信号通路的活性显著增加,表明与增殖和生长负相关的细胞生物学过程如凋亡和坏死在NeuroD1过表达细胞中得到促进,这与先前观察到的NeuroD1过表达抑制胰腺癌细胞增殖的结果一致。另一个重要发现是,识别出的差异表达基因在神经元相关通路中高度富集,如神经系统发育、GABA能突触和树突棘。这些发现提示NeuroD1过表达可能诱导胰腺癌细胞转分化为神经元。构建了神经发育通路的差异表达基因热图,确认了NeuroD1是该通路中的关键基因,同时也证实了NeuroD1过表达细胞已成功建立。
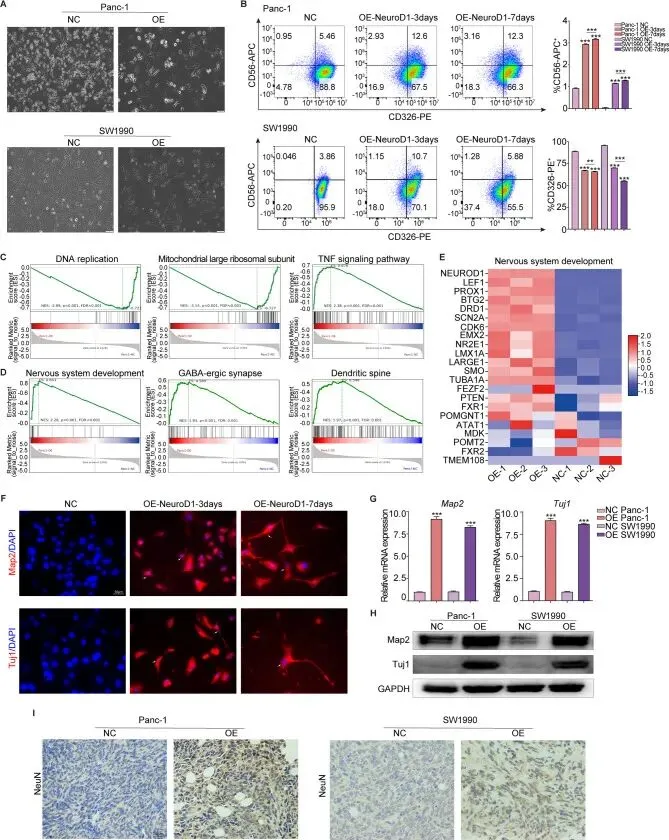 微管相关蛋白2、III类β-微管蛋白和神经元核蛋白是神经元中的重要蛋白标志物。Map2和Tuj1是关键的神经元细胞骨架蛋白,Tuj1常被用作早期神经元分化的标志物,NeuN则常用作成熟分化神经元的标志物。分选CD56阳性的NeuroD1过表达细胞,通过Map2和Tuj1染色进行免疫荧光染色以观察其细胞骨架形态。结果显示,与对照细胞相比,NeuroD1过表达后第3天部分细胞即呈现出神经元样形态,至第7天这些细胞显示出明显的神经元形态。这些发现支持NeuroD1确实诱导胰腺癌细胞转分化为神经元,并表明随着持续的NeuroD1过表达,转分化程度随时间增加。此外,通过RT-qPCR和Western Blotting实验证实,在NeuroD1过表达的胰腺癌细胞中,Map2和Tuj1的基因和蛋白水平表达均上调。同时,对先前体内实验收集的小鼠皮下肿瘤组织进行NeuN免疫组化染色,发现对照组织中几乎没有NeuN阳性细胞,而NeuroD1过表达细胞中存在显著比例的NeuN阳性细胞,表明部分胰腺癌细胞已转分化为成熟神经元样细胞。此外,通过流式细胞术分选CD56阳性和CD326阳性两个不同的细胞群体,并用这些分选的细胞群体在小鼠中建立皮下异种移植肿瘤,结果只有CD326阳性细胞形成肿瘤,而CD56阳性组未能形成肿瘤。综上所述,这些结果表明NeuroD1促进胰腺癌细胞转分化为神经元样细胞。
微管相关蛋白2、III类β-微管蛋白和神经元核蛋白是神经元中的重要蛋白标志物。Map2和Tuj1是关键的神经元细胞骨架蛋白,Tuj1常被用作早期神经元分化的标志物,NeuN则常用作成熟分化神经元的标志物。分选CD56阳性的NeuroD1过表达细胞,通过Map2和Tuj1染色进行免疫荧光染色以观察其细胞骨架形态。结果显示,与对照细胞相比,NeuroD1过表达后第3天部分细胞即呈现出神经元样形态,至第7天这些细胞显示出明显的神经元形态。这些发现支持NeuroD1确实诱导胰腺癌细胞转分化为神经元,并表明随着持续的NeuroD1过表达,转分化程度随时间增加。此外,通过RT-qPCR和Western Blotting实验证实,在NeuroD1过表达的胰腺癌细胞中,Map2和Tuj1的基因和蛋白水平表达均上调。同时,对先前体内实验收集的小鼠皮下肿瘤组织进行NeuN免疫组化染色,发现对照组织中几乎没有NeuN阳性细胞,而NeuroD1过表达细胞中存在显著比例的NeuN阳性细胞,表明部分胰腺癌细胞已转分化为成熟神经元样细胞。此外,通过流式细胞术分选CD56阳性和CD326阳性两个不同的细胞群体,并用这些分选的细胞群体在小鼠中建立皮下异种移植肿瘤,结果只有CD326阳性细胞形成肿瘤,而CD56阳性组未能形成肿瘤。综上所述,这些结果表明NeuroD1促进胰腺癌细胞转分化为神经元样细胞。Neuropathiazol上调胰腺癌中NeuroD1的表达
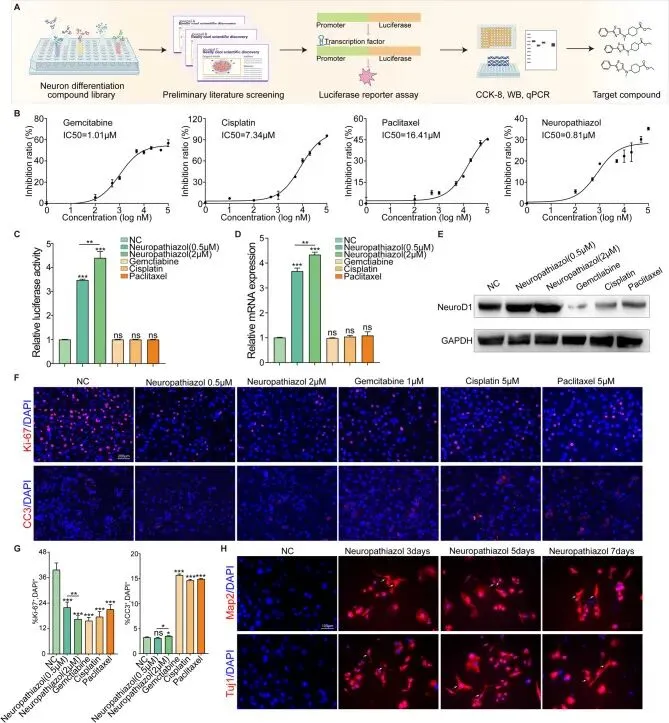
NeuroD1可诱导胰腺癌细胞从恶性表型向良性神经元表型转化,因此筛选调控和诱导NeuroD1表达的药物可能有助于胰腺癌的治疗。作者先前的研究已证明NeuroD1在髓母细胞瘤中的表达可通过表观遗传机制调控。然而,可用于调节此类表观遗传改变的H3K27甲基化抑制剂常伴有显著的毒性和副作用。鉴于这些局限性,有必要筛选更有效且毒性更低的药物,以有效调控胰腺癌中NeuroD1的特异性表达。为实现这一目标,进行了全面的药物筛选实验,同时使用EZH2抑制剂UNC1999作为对照,评估其对胰腺癌细胞中NeuroD1表达的影响。结果显示UNC1999不影响胰腺癌中NeuroD1的表达水平。
首先,基于与神经元分化相关的310种化合物库,初步识别出60种可能诱导NeuroD1表达的化合物。通过NeuroD1启动子荧光素酶报告实验,进一步识别出17种调控NeuroD1基因表达的化合物。最后,通过CCK-8实验评估这些化合物对细胞增殖的影响,并通过qPCR和Western Blotting验证NeuroD1的表达。最终鉴定出Neuropathiazol是一种能够诱导NeuroD1表达上调且具有低细胞毒性作用的小分子化合物。图示展示了药物筛选流程示意图,表格提供了NeuroD1的启动子序列,补充图显示了Panc-1和SW1990细胞经神经元分化化合物库中多种化合物处理后的NeuroD1启动子荧光素酶报告实验、CCK-8实验和qPCR实验结果。
通过CCK-8实验测定了Neuropathiazol及胰腺癌常用一线化疗药物吉西他滨、顺铂和紫杉醇的半数抑制浓度。结果显示,Neuropathiazol在抑制胰腺癌细胞增殖方面高度有效,甚至表现出优于传统一线化疗药物的抗肿瘤活性。此外,在特定浓度范围内,Neuropathiazol不损害正常胰腺上皮细胞的增殖能力。用PBS、不同浓度Neuropathiazol及化疗药物处理Panc-1细胞48小时,随后测量相对荧光素酶活性以评估NeuroD1启动子活性,结果显示常用一线化疗药物不影响Panc-1细胞中NeuroD1启动子活性,而Neuropathiazol处理则导致NeuroD1启动子活性呈浓度依赖性显著增加。通过qPCR和Western blotting检测Neuropathiazol及各种化疗药物处理后Panc-1细胞中NeuroD1在基因和蛋白水平上的表达变化,发现吉西他滨、顺铂和紫杉醇不诱导NeuroD1表达变化,而Neuropathiazol处理则引起NeuroD1表达呈浓度依赖性上调。这些结果表明,Neuropathiazol在胰腺癌细胞中诱导NeuroD1表达上调,且NeuroD1在体外展现出强效抗肿瘤活性。为深入探究Neuropathiazol的抗肿瘤活性,用上述浓度的各种药物处理Panc-1细胞48小时,并进行Ki-67和CC3免疫荧光染色以观察细胞增殖和凋亡。Ki-67蛋白免疫荧光染色显示,与对照组相比,Neuropathiazol或化疗药物治疗均能抑制细胞增殖,且Neuropathiazol浓度越高,增殖抑制作用越明显。CC3蛋白免疫荧光染色显示,与对照组相比,化疗药物显著增加了细胞凋亡,而Neuropathiazol仅引起轻微的凋亡增加或无变化。由此可得出结论:Neuropathiazol在体外具有有效的抗肿瘤活性。随后,在Neuropathiazol处理开始后的不同时间点进行Map2和Tuj1免疫荧光染色以观察细胞骨架形态。结果显示,Neuropathiazol处理诱导胰腺癌细胞形态转化为呈现神经元表型的细胞,随着处理时间的延长,这些细胞逐渐演变为形态类似于成熟神经元的细胞。基于上述发现,Neuropathiazol被鉴定为一种能够诱导并调控NeuroD1表达的药物,它上调胰腺癌细胞中NeuroD1的表达,并诱导胰腺癌细胞转分化为神经元。Neuropathiazol在体内抑制胰腺癌生长并诱导神经转分化
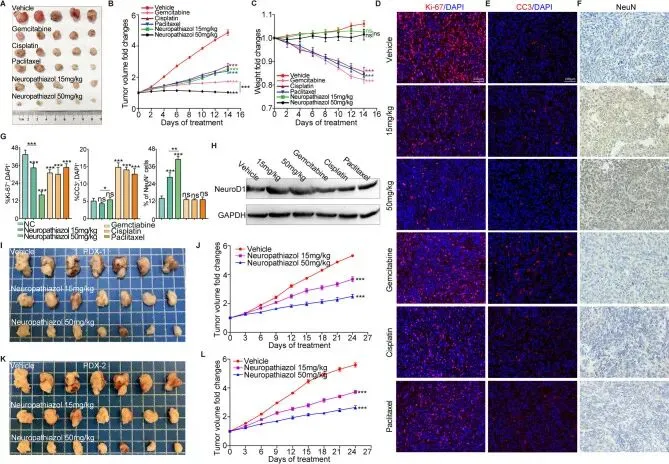
通过一系列体内实验进一步探索了Neuropathiazol的抗肿瘤功能。使用Panc-1细胞构建小鼠皮下肿瘤模型,当肿瘤体积接近200立方毫米时,将小鼠随机分为六组,分别接受不同剂量Neuropathiazol、吉西他滨、顺铂或紫杉醇治疗,对照组给予溶剂对照,所有药物每两天腹腔注射一次,持续两周。结果显示,Neuropathiazol及三种化疗药物均能抑制皮下肿瘤的体内生长,其中15 mg/kg Neuropathiazol的抑瘤程度与化疗药物相似,而50 mg/kg Neuropathiazol则更显著地抑制了肿瘤生长。在给药期间监测动物体重发现,化疗药物导致显著的体重下降,而Neuropathiazol治疗未引起明显的体重变化,提示Neuropathiazol治疗不引起减重这一副作用。对皮下肿瘤进行Ki-67和CC3蛋白免疫荧光染色,结果显示Neuropathiazol和化疗药物均能显著抑制体内胰腺癌细胞的增殖,且较高浓度的Neuropathiazol产生更明显的抗增殖效果;同时,化疗药物促进胰腺癌细胞凋亡,而Neuropathiazol短期内未显著增加凋亡。皮下肿瘤中NeuN蛋白的免疫组化染色显示,Neuropathiazol治疗后NeuN阳性细胞比例显著增加,且呈浓度依赖性。Western blotting检测皮下肿瘤中NeuroD1表达的结果显示,Neuropathiazol上调肿瘤组织中NeuroD1的表达,而化疗药物则无此作用。这些发现表明Neuropathiazol以浓度依赖性方式诱导胰腺癌细胞转分化为神经元,而所测试的化疗药物不诱导相应的细胞转分化。
随后从两例胰腺癌患者收集胰腺导管腺癌组织,皮下植入NSG小鼠以构建患者来源异种移植模型。当肿瘤体积达到约200立方毫米时,将荷瘤小鼠随机分为三组,分别给予溶剂对照、15 mg/kg Neuropathiazol或50 mg/kg Neuropathiazol治疗,每三天腹腔注射一次,治疗期间监测肿瘤体积变化。药物治疗后第24天收集肿瘤并拍照。结果显示,Neuropathiazol显著抑制了小鼠体内患者来源皮下肿瘤的进展,且其抑制效应随Neuropathiazol浓度增加而增强。这些体内实验清楚地表明,Neuropathiazol在体内有效抑制胰腺癌的进展。在此基础上,进一步研究了Neuropathiazol与化疗药物联合的治疗效果。使用Panc-1细胞建立小鼠皮下肿瘤模型,当肿瘤体积接近200立方毫米时,将小鼠随机分为四组,分别给予溶剂对照、吉西他滨、Neuropathiazol以及Neuropathiazol联合吉西他滨治疗,每两天腹腔注射一次,持续两周。结果显示,与吉西他滨相比,Neuropathiazol表现出更显著的体内肿瘤生长抑制能力;值得注意的是,Neuropathiazol与吉西他滨的联合进一步增强了皮下肿瘤生长的抑制效果。皮下肿瘤中Ki-67蛋白的免疫荧光染色也显示,Neuropathiazol与吉西他滨联合进一步抑制了胰腺癌细胞在体内的增殖。这些发现提示转分化诱导剂Neuropathiazol与化疗药物的联合可能通过进一步清除残留肿瘤细胞而发挥更显著的肿瘤抑制作用。Neuropathiazol结合MET蛋白并抑制其磷酸化
接下来,旨在阐明Neuropathiazol的作用靶点及其诱导NeuroD1表达上调的具体机制。
利用已知的Neuropathiazol二维结构及MOE软件生成该化合物的初步三维结构,在Amber10-EHT力场下通过Wash功能添加氢原子,并在刚性溶剂水条件下以0.1 kcal/mol/Å的最佳梯度均方根截断值对三维结构进行能量最小化。随后利用PharmMapper、SwissTargetPrediction和GalaxySagittarius方法预测Neuropathiazol的潜在靶点,这些方法共揭示Neuropathiazol有175个可能靶点。同时,使用GeneCards v5.15以“胰腺癌”为搜索词收集已知的胰腺癌相关靶点,通过设定评分阈值大于30,识别出336个与胰腺癌强相关的靶点。对数据库筛选获得的175个靶点与人工筛选识别的336个靶点进行交集分析,识别出15个两种方法均识别且经MOE软件确认能与Neuropathiazol对接的靶点。使用MOE软件评估蛋白质与Neuropathiazol之间的相互作用能,评分值以S值表示,S值越低表示两者亲和力越强。在识别的15个潜在靶点中,确定MET蛋白是唯一的膜蛋白,且似乎是Neuropathiazol最可能的靶点。
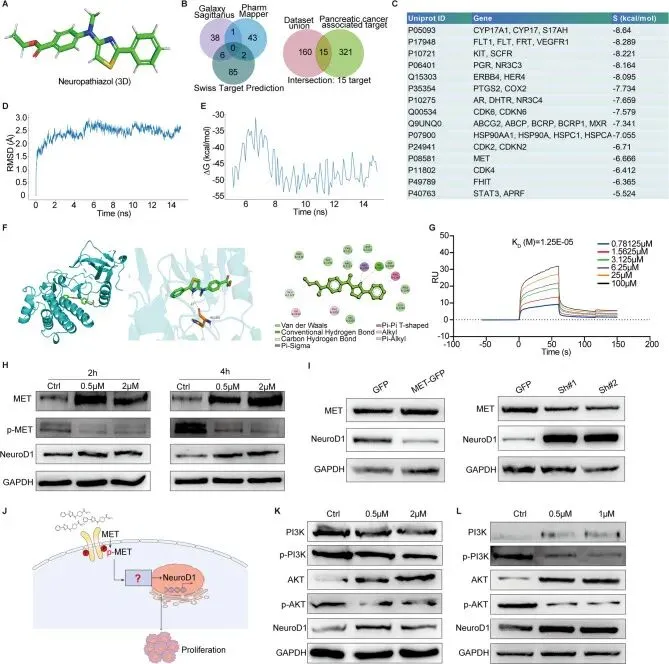 为探究Neuropathiazol是否稳定结合MET,进行了15纳秒分子动力学模拟。模拟结果的RMSD分析显示系统在2纳秒后开始收敛,结构在大约5纳秒时达到平衡,整体RMSD值在2.5埃左右波动。随后选取5-15纳秒时间范围内的结构进行MM-GBSA结合自由能分析,结果显示结合自由能在-50 kcal/mol左右稳定,平均值为-46.60 kcal/mol,其中8-15纳秒的平均结合自由能为-48.96 kcal/mol,表明两者之间存在强相互作用。通过计算机模拟Neuropathiazol-MET蛋白复合物的构象,发现Neuropathiazol通过与His-1202形成氢键与MET蛋白结合,具体结合位于MET蛋白氨基酸残基1127至1260之间,该序列位于MET蛋白的磷酸化区域内。为进一步验证这种相互作用,采用表面等离子共振技术测定MET蛋白与Neuropathiazol之间的亲和力,结果显示解离常数为12.5e-5,表明Neuropathiazol与MET蛋白之间具有非常强的亲和力。这些发现表明MET蛋白很可能是Neuropathiazol的靶点,两者具有强亲和力,Neuropathiazol可能通过结合MET蛋白并调节其磷酸化而发挥作用。用不同浓度Neuropathiazol处理Panc-1细胞2-4小时,以PBS为对照,收集蛋白并通过Western Blotting评估MET和磷酸化MET的表达水平,观察到Neuropathiazol抑制Panc-1细胞中MET蛋白的磷酸化,随后NeuroD1表达上调。通过构建MET过表达和MET敲低的Panc-1细胞,发现MET过表达时NeuroD1表达下调,而MET敲低时NeuroD1表达增加。这些发现共同支持Neuropathiazol通过干扰MET蛋白磷酸化进而调节下游信号通路上调NeuroD1表达的结论。文献回顾表明,MET磷酸化可激活PI3K-AKT信号通路,因此通过Western Blotting检测Neuropathiazol处理后细胞中PI3K、磷酸化PI3K、AKT、磷酸化AKT和NeuroD1的表达水平,发现Neuropathiazol处理抑制了PI3K和AKT的磷酸化,同时增加了NeuroD1的表达。类似地,使用PI3K化学抑制剂LY294002阻断PI3K-AKT信号通路的激活也诱导了NeuroD1表达的上调。这些结果表明Neuropathiazol结合MET蛋白,抑制其磷酸化,从而抑制下游PI3K-AKT信号通路的激活,最终导致NeuroD1表达上调。
为探究Neuropathiazol是否稳定结合MET,进行了15纳秒分子动力学模拟。模拟结果的RMSD分析显示系统在2纳秒后开始收敛,结构在大约5纳秒时达到平衡,整体RMSD值在2.5埃左右波动。随后选取5-15纳秒时间范围内的结构进行MM-GBSA结合自由能分析,结果显示结合自由能在-50 kcal/mol左右稳定,平均值为-46.60 kcal/mol,其中8-15纳秒的平均结合自由能为-48.96 kcal/mol,表明两者之间存在强相互作用。通过计算机模拟Neuropathiazol-MET蛋白复合物的构象,发现Neuropathiazol通过与His-1202形成氢键与MET蛋白结合,具体结合位于MET蛋白氨基酸残基1127至1260之间,该序列位于MET蛋白的磷酸化区域内。为进一步验证这种相互作用,采用表面等离子共振技术测定MET蛋白与Neuropathiazol之间的亲和力,结果显示解离常数为12.5e-5,表明Neuropathiazol与MET蛋白之间具有非常强的亲和力。这些发现表明MET蛋白很可能是Neuropathiazol的靶点,两者具有强亲和力,Neuropathiazol可能通过结合MET蛋白并调节其磷酸化而发挥作用。用不同浓度Neuropathiazol处理Panc-1细胞2-4小时,以PBS为对照,收集蛋白并通过Western Blotting评估MET和磷酸化MET的表达水平,观察到Neuropathiazol抑制Panc-1细胞中MET蛋白的磷酸化,随后NeuroD1表达上调。通过构建MET过表达和MET敲低的Panc-1细胞,发现MET过表达时NeuroD1表达下调,而MET敲低时NeuroD1表达增加。这些发现共同支持Neuropathiazol通过干扰MET蛋白磷酸化进而调节下游信号通路上调NeuroD1表达的结论。文献回顾表明,MET磷酸化可激活PI3K-AKT信号通路,因此通过Western Blotting检测Neuropathiazol处理后细胞中PI3K、磷酸化PI3K、AKT、磷酸化AKT和NeuroD1的表达水平,发现Neuropathiazol处理抑制了PI3K和AKT的磷酸化,同时增加了NeuroD1的表达。类似地,使用PI3K化学抑制剂LY294002阻断PI3K-AKT信号通路的激活也诱导了NeuroD1表达的上调。这些结果表明Neuropathiazol结合MET蛋白,抑制其磷酸化,从而抑制下游PI3K-AKT信号通路的激活,最终导致NeuroD1表达上调。NeuroD1与Neurog3相互作用抑制胰腺癌细胞增殖并诱导转分化
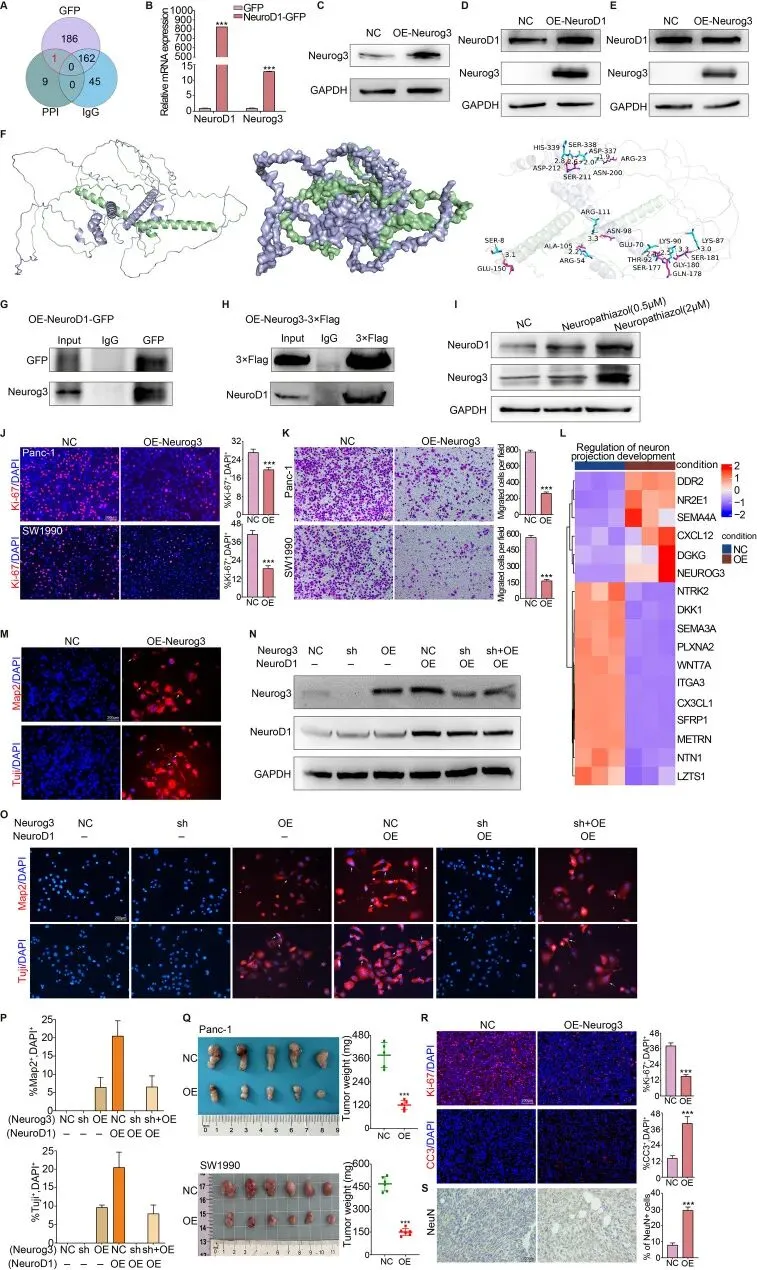
接下来寻找与NeuroD1相互作用的下游蛋白。构建了GFP标记的NeuroD1过表达Panc-1细胞,并进行Co-IP联合质谱分析。维恩图包含三部分数据:Co-IP实验中与正常IgG相互作用的蛋白、Co-IP实验中与NeuroD1-GFP相互作用的蛋白以及蛋白质-蛋白质相互作用网络数据库中显示与NeuroD1相互作用的蛋白。通过综合分析,最终鉴定出Neurog3是与NeuroD1相互作用的下游蛋白。Neurog3是隶属于碱性螺旋-环-螺旋转录因子家族的关键转录因子,先前已被证明能激活内分泌前体细胞中的基因转录,在胰腺和肠道内分泌细胞谱系形成中起关键作用。为探究NeuroD1与Neurog3的关系,首先在NeuroD1过表达Panc-1细胞中通过RT-qPCR评估两者的表达水平,发现随着NeuroD1表达增加,Neurog3表达相应上调。随后建立Neurog3过表达细胞系并通过Western Blotting确认构建成功,进一步分析这些过表达细胞系中NeuroD1和Neurog3的表达,结果显示NeuroD1过表达持续导致Neurog3表达水平升高,但Neurog3过表达并不反过来导致NeuroD1上调。这些发现表明NeuroD1与Neurog3之间可能存在相互作用或调控关系,其中NeuroD1调节Neurog3的表达。
为进一步阐明NeuroD1与Neurog3蛋白之间的相互作用,利用UniProtKB数据库进行了蛋白-蛋白对接分析。结果显示NeuroD1与Neurog3之间具有强结合亲和力,结合自由能为-16.6 kcal/mol。图示展示了整体对接构型,其中绿色表示Neurog3蛋白,紫色表示NeuroD1蛋白。表面模型显示NeuroD1与Neurog3蛋白之间的相互作用通过在其接触表面形成广泛的氢键而稳定,从而增加了蛋白复合物的整体稳定性。对关键氨基酸残基的详细分析显示,关键残基之间的多个接触点形成氢键,进一步增强了结合的稳定性。总体而言,蛋白-蛋白对接分析证明NeuroD1与Neurog3之间存在强大的表面相互作用,具有高度稳定的结合能。这种强而稳定的结合提示这两种蛋白之间的相互作用达到了最佳状态,并为它们潜在的功能关系提供了见解。为证实NeuroD1与Neurog3之间的物理相互作用,使用GFP标记的NeuroD1过表达Panc-1细胞和3×Flag标记的Neurog3过表达Panc-1细胞进行Co-IP实验。结果显示,Neurog3可被NeuroD1-GFP特异性抗体有效沉淀,而NeuroD1也可被Neurog3-3×Flag特异性抗体成功沉淀。这些发现为NeuroD1与Neurog3之间的直接相互作用提供了进一步证据。此外,Neuropathiazol处理导致Panc-1细胞中NeuroD1和Neurog3水平同时上调。随后研究了Neurog3过表达对Panc-1和SW1990细胞增殖和迁移能力的影响,发现Neurog3过表达显著抑制了胰腺癌细胞的增殖和迁移。为进一步阐明这些效应的机制,对对照Panc-1细胞和Neurog3过表达Panc-1细胞进行RNA测序并分析差异表达基因,结果显示差异表达基因在与神经元投射发育调控相关的信号通路中显著富集。这些发现提示Neurog3可能在促进胰腺癌细胞向神经元样表型转化中发挥作用。与此一致,对Neurog3过表达细胞进行Map2和Tuj1免疫荧光染色显示,Neurog3上调了胰腺癌细胞向神经元样表型的转分化。使用Panc-1细胞建立了对照、Neurog3敲低和Neurog3过表达细胞系,同时构建了NeuroD1过表达Panc-1细胞的双调控模型,包含对照、Neurog3敲低和Neurog3挽救组。Map2和Tuj1免疫荧光染色显示,在NeuroD1低表达的胰腺癌细胞中,无论是对照组还是Neurog3敲低组均未表现出向神经元样表型的转变,而Neurog3过表达部分诱导了这一转变。在NeuroD1高表达的细胞中,Neurog3敲低抑制了神经元样转变,而恢复Neurog3表达可挽救这种抑制。在六个细胞组中,高表达NeuroD1且未敲低Neurog3的Panc-1细胞表现出最高比例的神经元样表型转变细胞。各组中转分化细胞比例的量化显示,NeuroD1的转分化诱导效应依赖于Neurog3。然而,就诱导效力而言,NeuroD1在诱导胰腺癌细胞转分化为神经元样表型方面表现出显著高于Neurog3的能力。使用Panc-1和SW1990细胞在小鼠中建立皮下肿瘤模型,用于研究Neurog3过表达对体内胰腺癌进展的影响,发现Neurog3过表达显著抑制了胰腺癌的体内进展。收集皮下肿瘤组织并进行Ki-67、CC3和NeuN蛋白的免疫荧光及免疫组化染色,结果显示Neurog3过表达显著降低了胰腺癌细胞的增殖能力,同时显著增加了细胞凋亡。表达神经元标志物的细胞比例也显著增加,表明Neurog3诱导胰腺癌细胞向神经元样细胞的表型转化。综上所述,研究结果表明NeuroD1与Neurog3相互作用,并在体外和体内抑制胰腺癌细胞的增殖,同时诱导胰腺癌细胞转分化为神经元样表型的细胞。MET-NeuroD1-Neurog3轴抑制胰腺癌的肿瘤进展
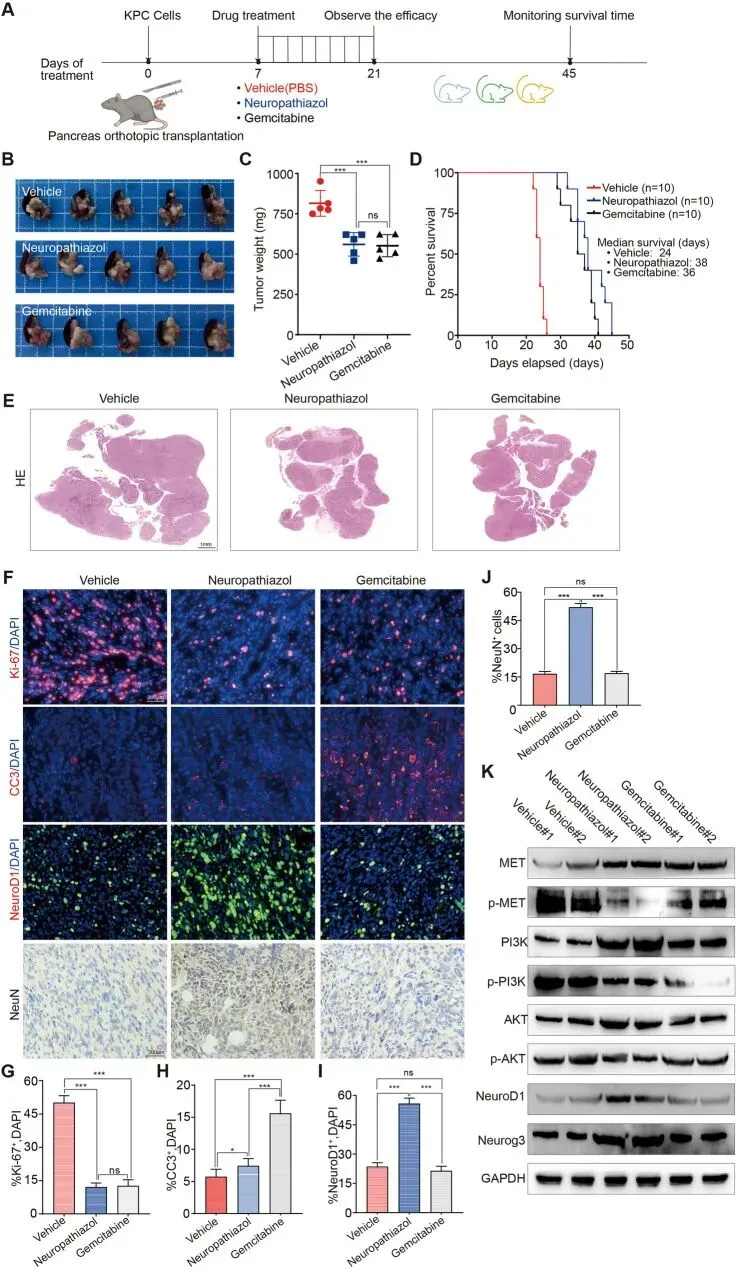
从KPC转基因小鼠的自发肿瘤中分离胰腺癌细胞,建立原位胰腺移植肿瘤模型。将小鼠随机分为三组,在肿瘤植入后第7天开始,每两天腹腔注射溶剂对照、Neuropathiazol或吉西他滨,持续两周。两周后,收获原位胰腺肿瘤并拍照称重;另一批荷瘤小鼠接受持续给药直至死亡。实验流程图显示,Neuropathiazol治疗显著抑制了胰腺肿瘤生长并延长了小鼠的生存时间,显示出与吉西他滨相当的治疗效果。
在Neuropathiazol给药后的多个时间点采集小鼠血浆样本,通过高效液相色谱-串联质谱法对Neuropathiazol浓度进行定量。结果显示Neuropathiazol在给药后0.083小时即达到血浆峰值浓度,随后浓度逐渐下降,至8小时几乎检测不到。为研究其组织分布,在峰值时间点及后续两个时间点定量检测小鼠血浆、心脏、肝脏、肾脏、肺和脾脏中的Neuropathiazol浓度,结果显示Neuropathiazol被组织良好吸收,峰值时间点后其浓度逐渐下降。详细的药代动力学数据、体内剂量-反应曲线和组织分布数据已呈现。药代动力学数据表明Neuropathiazol起效迅速,且易于从体内清除而不蓄积。此外,治疗期间对各组小鼠体重变化的持续监测显示,Neuropathiazol治疗未显著改变体重,而吉西他滨治疗导致显著体重下降。收集各组小鼠的心脏、肝脏、肾脏、肺和脾脏组织进行H&E染色,结果显示Neuropathiazol未造成显著的器官损伤。综上所述,这些发现表明Neuropathiazol起效迅速,易于从体内清除而不蓄积,且具有低药物毒性和良好的安全性。肿瘤组织的H&E染色证实了原位胰腺移植肿瘤的成功建立。对肿瘤组织进行Ki-67、CC3和NeuroD1的免疫荧光染色,结果显示Neuropathiazol和吉西他滨均显著抑制了胰腺癌细胞的增殖并促进了细胞凋亡;Neuropathiazol治疗还上调了胰腺癌细胞中NeuroD1的表达。NeuN蛋白的免疫组化染色显示,Neuropathiazol增加了胰腺癌组织中NeuN阳性细胞的比例,表明Neuropathiazol诱导部分胰腺癌细胞转分化为神经元样细胞,而吉西他滨治疗后未观察到这一效应。基于这些结果,提出吉西他滨通过细胞毒性发挥抗肿瘤作用导致胰腺癌细胞死亡,而Neuropathiazol通过使胰腺癌细胞向更分化的神经元样良性表型转化来抑制肿瘤进展。通过Western Blotting检测Neuropathiazol或吉西他滨处理的肿瘤组织中MET、PI3K和AKT蛋白的表达及磷酸化水平,结果显示Neuropathiazol抑制了MET蛋白的磷酸化及PI3K-AKT信号通路的活性,从而增加NeuroD1和Neurog3的表达水平,而吉西他滨不改变这些蛋白的表达水平。综上所述,研究结果证明Neuropathiazol通过MET-NeuroD1-Neurog3轴抑制胰腺癌进展。讨论:
本研究首次将NeuroD1介导的神经元转分化机制引入胰腺癌,发现NeuroD1可通过Neurog3诱导肿瘤细胞转分化为神经元样细胞,从而抑制其恶性表型。通过药物筛选,鉴定出小分子化合物Neuropathiazol,其通过与MET蛋白结合抑制PI3K-AKT信号通路,特异性上调NeuroD1表达。体内实验显示,Neuropathiazol联合化疗药物可显著缩小肿瘤体积,并激活TNFα信号通路及促凋亡机制。研究还提示转分化可能重塑肿瘤免疫微环境,增强免疫识别。尽管该策略展现出转化潜力,但仍存在局限:肿瘤细胞与神经微环境的交互可能存在潜在风险;转分化后神经元样细胞的长期命运及表型稳定性尚不明确;需结合谱系示踪、单细胞测序和空间组学技术进一步验证其机制与安全性。未来应系统开展靶点验证、剂量优化和长期监测,探索与免疫检查点抑制剂等联合策略,为临床转化奠定基础。关注我 定期分享最新sci热点思路
定期分享最新sci热点思路 夜雨聆风
夜雨聆风