 夜雨聆风
夜雨聆风我们或许都有过这样的经历:站在超市的冷藏柜前,面对琳琅满目的“益生菌酸奶”或保健品,暗自期待它们能拯救自己脆弱的肠胃、缓解代谢焦虑或是唤醒疲惫的免疫系统。然而,这些传统的“第一代”益生菌在进入人体后,大多只是匆匆过客,难以真正驻留并发挥持久的、可预测的临床疗效。
科学界对这一“疗效悖论”的探索从未停止。2026年5月8日,国际顶级学术期刊《Nature Reviews Microbiology》发表了一篇具有里程碑意义的重磅综述《Next-generation probiotics: an outlook into current applications and future developments》。来自魏茨曼科学研究所和德国癌症研究中心的Eran Elinav教授团队,全面描绘了从传统食品衍生菌株向“下一代益生菌”(NGP,又称精准益生菌)跨越的宏伟蓝图。研究不仅深刻剖析了活体细菌疗法在定植抵抗、宿主内进化及安全性等方面的核心挑战,更以前所未有的视野,展示了人工智能与计算生物学如何重塑精准微生态疗法的未来。
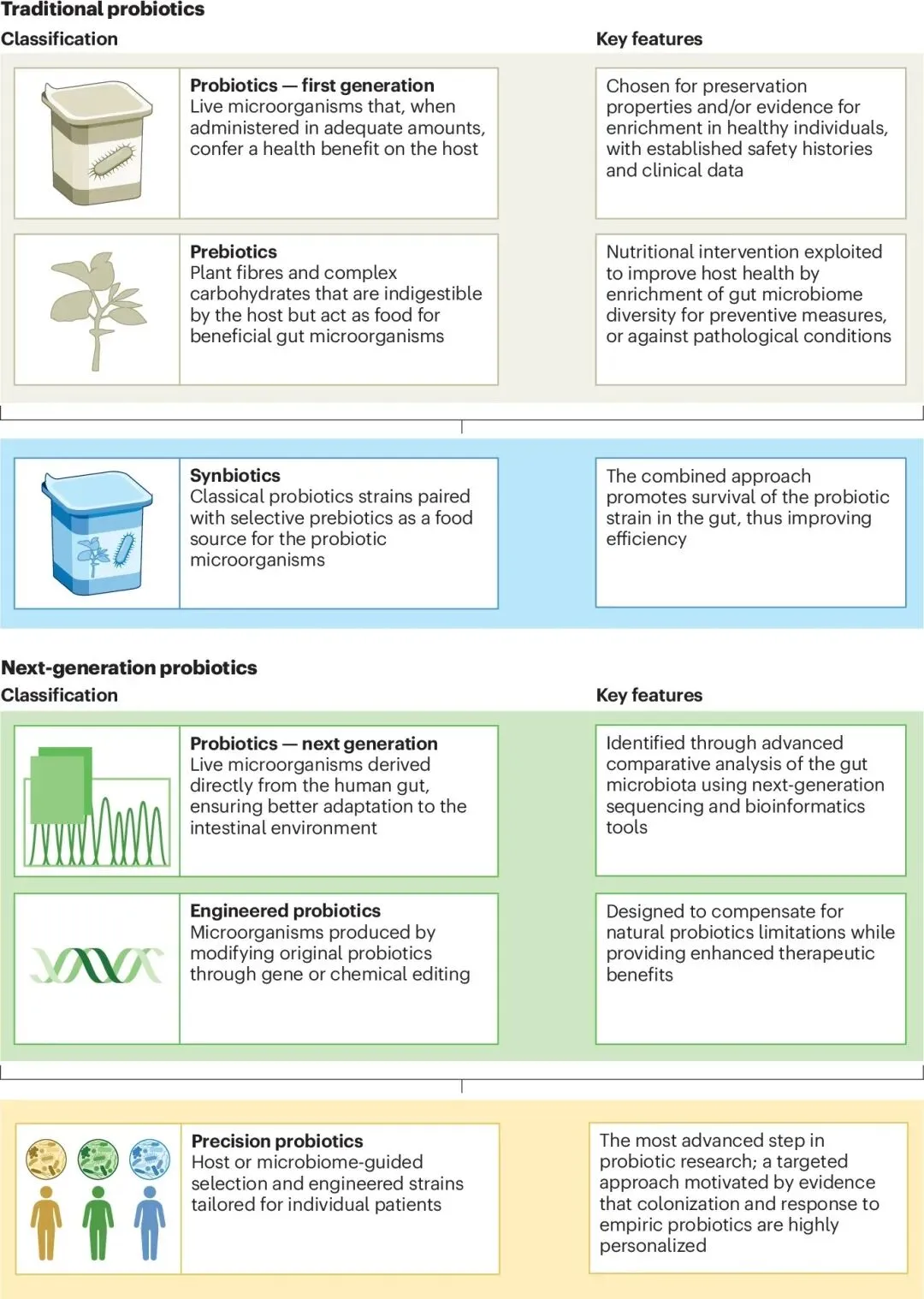
图1 下一代益生菌疗法的演进
盲盒式补充的终结,传统益生菌的定植壁垒与疗效悖论
长期以来,以乳酸杆菌和双歧杆菌为代表的“第一代益生菌”占据了绝大部分市场。它们多源自发酵食品,凭借悠久的安全食用历史,被广泛作为膳食补充剂销售,从而巧妙避开了现代药物开发所需的严格多期临床试验与审批流程。然而,这种“一刀切”的盲盒式补充策略,完全忽视了宿主遗传背景、免疫状态以及基线肠道菌群的巨大个体差异。临床实践和严谨的科学研究无情地揭示:许多备受推崇的传统益生菌株在进入人体后,根本无法克服复杂的生态壁垒,最终只能随粪便排出,未能真正在肠道黏膜层实现稳定定植,其宣称的健康益处也因此饱受科学界的广泛质疑与激烈辩论[1]。
阻碍益生菌发挥疗效的最大拦路虎,是人体自身构建的“定植抵抗”机制。当外来菌株试图在胃肠道安营扎寨时,它们首先要经历极端物理化学环境的毒打——包括胃部极低的pH值、胆盐的强力洗刷以及消化酶的降解。即便侥幸存活,它们还要面对人体原籍肠道菌群构筑的坚不可摧的生态防线。常驻共生菌通过极其复杂的机制驱逐“外来户”:例如,大肠杆菌Nissle株会分泌微菌素等抗菌分子直接进行生化武器攻击;另一些共生菌则通过大量消耗核心营养物质,或分泌短链脂肪酸来酸化局部微环境,从根本上切断外来菌株的生存资源。此外,常驻真菌和细菌还能激活宿主的先天与适应性免疫反应,促使肠道上皮分泌抗菌肽,形成一道密不透风的免疫屏障[2]。
除了定植失败的无效性,传统益生菌在特定语境下的安全性盲区同样令人警醒。虽然对健康人群普遍安全,但在免疫力低下的特殊人群中,盲目使用传统益生菌曾引发过致命的系统性败血症。更具颠覆性的发现是,在抗生素治疗后服用传统益生菌,不仅未能如预期般加速肠道微生态的恢复,反而会引发长期的微生态失调,甚至在无意中扩大了肠道内的“抗生素耐药基因库”,导致携带万古霉素耐药基因的细菌异常扩增[3]。这些残酷的临床现实迫切呼唤一种全新的干预手段:它们必须具备高度的生态契合度、清晰的分子机制,并符合现代生物制药的严苛标准。
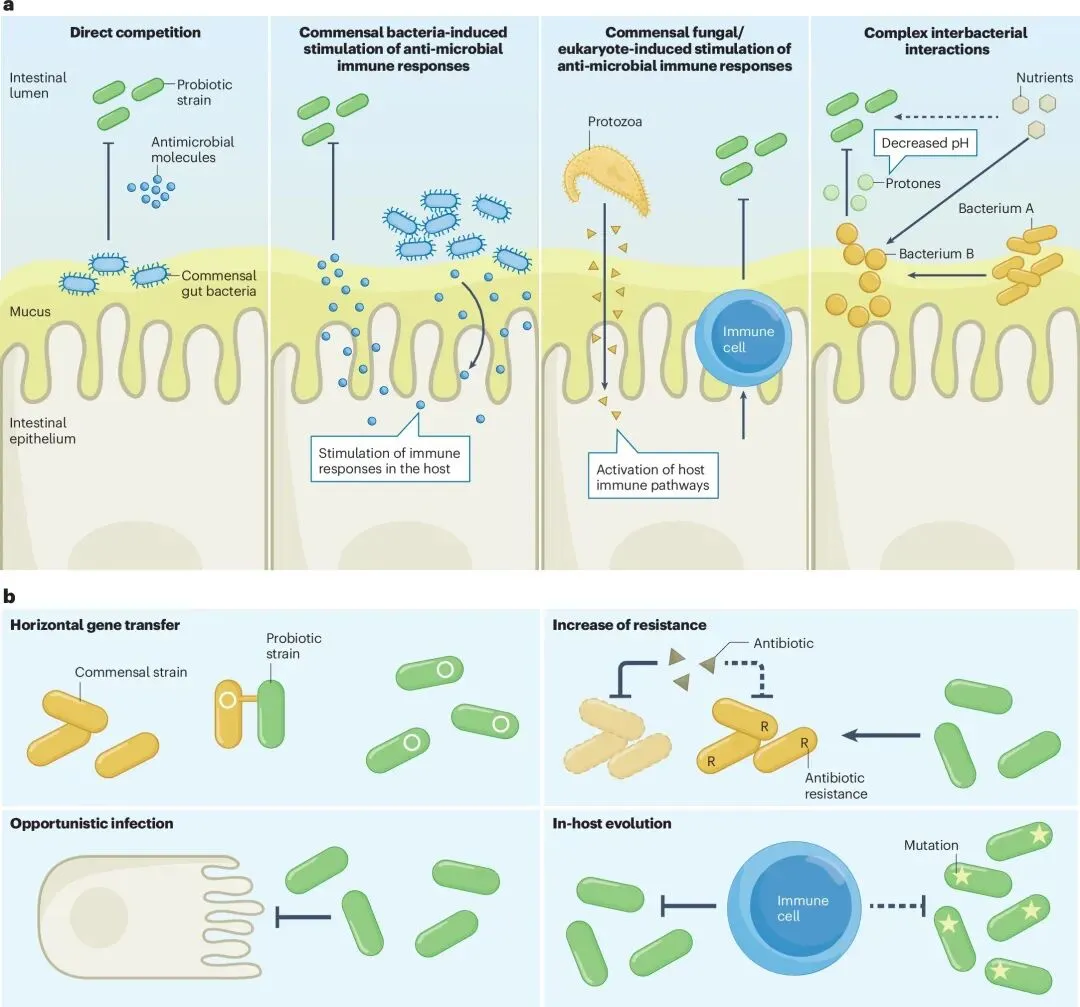
图2 NGP使用的挑战与风险
数据与算法的交汇,多组学解码与高分辨率代谢网络构建
与第一代益生菌多来源于食品不同,下一代益生菌的发现高度依赖于对人类肠道微生态的深度挖掘。研究人员通过宏基因组学、宏转录组学以及代谢组学等高通量测序技术,对海量健康人群与疾病队列的样本进行对比分析。这种“自下而上”的数据驱动策略,使得科学家能够精准锁定那些在健康状态下富集、在疾病状态下缺失的关键候选菌株。这些源自人类自身的“原住民”菌株,天生就具备更好的肠道适应性和生态位竞争力。然而,面对数以千计的菌株和海量的代谢产物,传统的统计学方法已捉襟见肘,人工智能与计算生物学的介入成为了破局的关键。
为了从浩如烟海的微生物数据中筛选出具有治疗潜力的菌株,研究团队引入了微生物群落规模的代谢网络模型。以最新发布的AGORA2数据库为例,该资源库包含了高达7,302个高质量、菌株水平的人类相关微生物基因组规模代谢重建[4]。通过这些精密的计算模型,科学家不再仅仅停留在“谁在那里”的物种丰度层面,而是深入到“它们在干什么”的功能层面。AGORA2甚至整合了98种常用处方药的代谢途径,能够基于个体的年龄、性别、体重指数和临床表型,精准预测个体化肠道菌群对药物的生物转化能力。这种基于约束的机制建模,为精准医疗提供了前所未有的高分辨率视角。
在代谢物产出预测方面,机器学习展现出了惊人的效率。一项前沿研究利用代谢网络模型和机器学习技术,旨在筛选高效产丁酸的微生物群落。研究人员首先为19种肠道分离菌株构建了基因组规模的代谢模型,随后在计算机中模拟了超过45万种合成菌群的组合,并训练回归模型来预测丁酸的产量[5]。这种计算机模拟筛选极大地缩小了候选范围,随后的湿实验验证高度吻合了算法的预测结果。这种将深度学习模式识别与生物学机制约束完美融合的策略,彻底改变了下一代益生菌的研发管线,使得从庞大的基因组库中快速定位目标代谢物产出菌株成为现实。
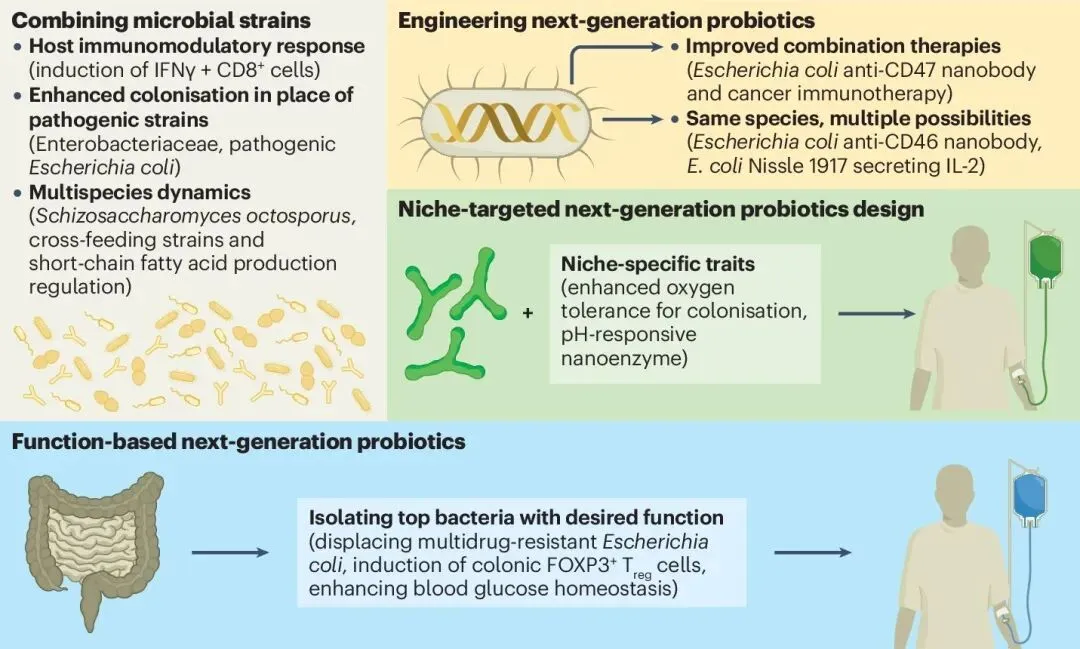
图3 NGP研发中的新兴策略
突破生物学极限,下一代益生菌的四大前沿设计策略
面对复杂的疾病机制与肠道生态,下一代益生菌的开发已经脱离了单一的“寻宝”模式,演化出四条极具颠覆性的前沿设计路径。首先是构建明确的多菌株甚至跨界合成菌群。面对根深蒂固的病原体,单一益生菌往往势单力薄。一项震撼学界的研究构建了一个包含119株人类衍生菌的庞大合成群落,通过反复的体内定植挑战,该“兵团”展现出了对致病性大肠杆菌的绝对定植抵抗力,并完美重塑了宿主的免疫与代谢表型[6]。在更具靶向性的设计中,科学家筛选出一个由18株共生菌组成的群落,它们能够通过对碳源葡萄糖酸盐的激烈竞争,精准剥夺多重耐药肠杆菌科的生存资源,在克罗恩病小鼠模型中实现了病原体的功能性去定植与炎症缓解。
其次是严格遵循“功能优先”的导向发现原则。科学家首先明确疾病治疗所需的特定生物学功能,再去浩瀚的菌群库中按图索骥。以嗜黏蛋白阿克曼菌(俗称AKK菌)为例,它最初因在健康瘦弱人群体内丰度显著高于肥胖人群而被关注。机制研究证实,其外膜蛋白能够特异性与宿主受体结合,强化肠道屏障并改善代谢稳态。如今,AKK菌已在人体临床试验中被证实能安全有效地改善超重志愿者的胰岛素敏感性[7]。同样,针对致命的移植物抗宿主病,研究人员直接基于体外短链脂肪酸的高产出能力,筛选出6种菌株组成鸡尾酒疗法,成功在小鼠模型中大幅提升了肠道代谢物浓度并显著提高了生存率。
当天然菌株无法同时满足定植与特定疗效的双重要求时,基因工程益生菌赋予了活体疗法无限可能。通过基因编辑,科学家将益生菌改造成能够在体内特定部位释放药物的智能可编程导弹。例如,研究人员将大肠杆菌Nissle 1917株进行改造,使其能够分泌具有免疫调节作用的白细胞介素-2,并为其穿上肠溶性聚合物外衣以抵抗胃酸。这种工程菌在口服后能精准抵达发炎的肠道释放免疫因子,显著促进了调节性T细胞的分化,在炎症性肠病模型中取得了惊艳的黏膜修复效果[8]。在肿瘤治疗中,一种被设计为能在肿瘤微环境中发生同步裂解的工程化大肠杆菌,能够精准释放抗CD47纳米抗体,不仅引发了强烈的肿瘤消退,与免疫检查点抑制剂联用时更激发了全身性的抗肿瘤免疫。
最后是微环境自适应的精准设计。共生菌具有独特的代谢适应性,利用这一特性,科学家正在设计能够精准响应特定病理微环境的益生菌。在结直肠癌的治疗探索中,研究人员将长双歧杆菌与一种pH响应型的过氧化物酶样纳米酶结合。这个精妙的系统只有在酸性的肿瘤微环境中才会被激活:温和的活性氧首先刺激益生菌代谢产酸,进一步触发纳米酶产生大量致命的活性氧以精准杀伤癌细胞[9]。这种将活体微生物的趋瘤性与纳米材料的物理化学开关完美结合的策略,展现了极高的肿瘤特异性和极低的全身毒性。与传统发酵食品来源、疗效高度个体化且缺乏精准靶点的第一代益生菌相比,下一代益生菌彻底完成了从“膳食补充剂”向机制明确、需经严格临床审批的“活体生物药”的华丽蜕变。
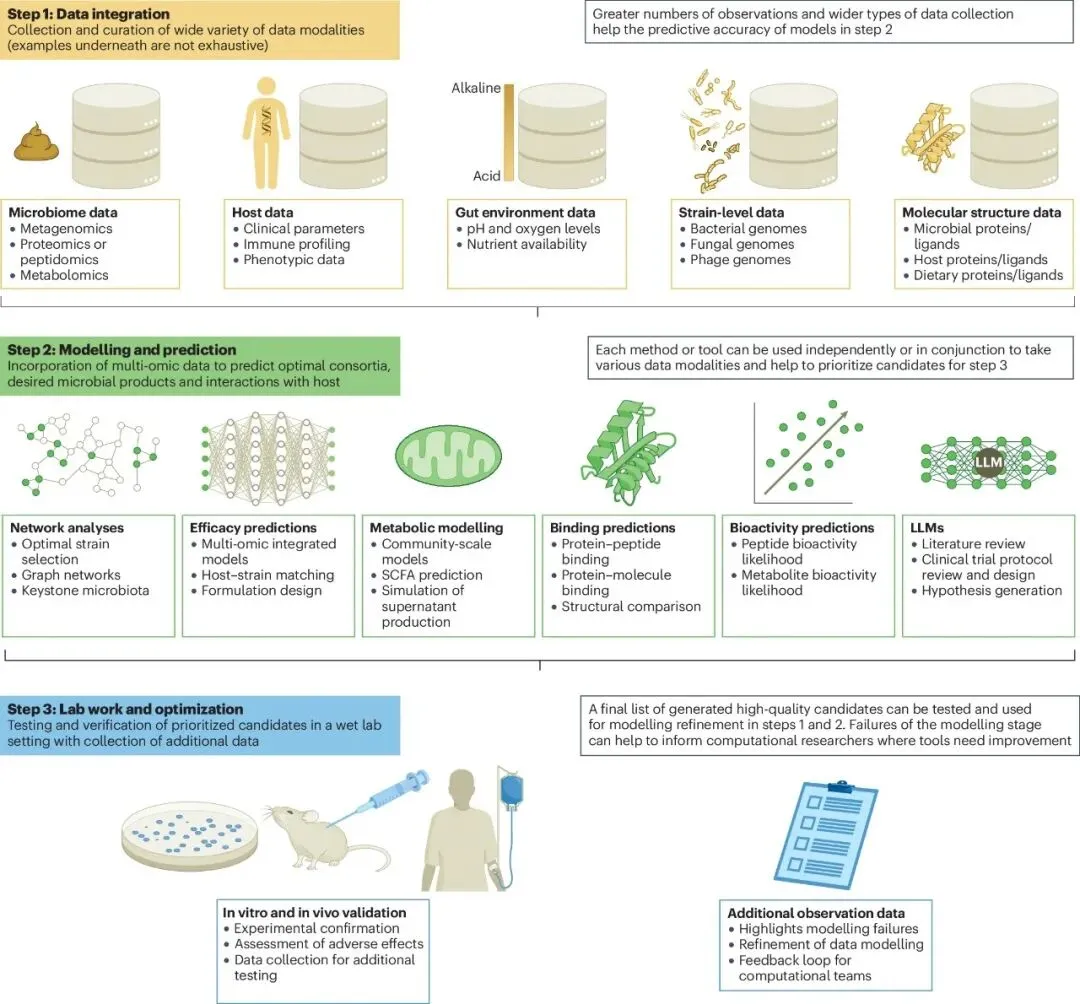
图4 下一代益生菌迭代设计与验证的概念框架
揭开微生态的“黑匣子”,跨界通讯与宿主内进化的双刃剑
下一代益生菌之所以能发挥奇效,关键在于其与宿主之间精妙的分子对话。传统益生菌时代的“黑匣子”正在被现代分子生物学层层拆解。研究揭示,益生菌并非简单地“占据地盘”,而是通过分泌海量的代谢物和活性蛋白,直接参与宿主的生理调控。例如,普拉梭菌能够分泌微生物抗炎分子,直接抑制宿主细胞的炎症通路并诱导抗炎因子的产生,成为治疗克罗恩病的明星候选菌[10]。此外,跨界通讯的发现更是拓宽了治疗视野。在泛癌免疫检查点抑制剂治疗队列中,科学家发现特定的真菌(如八孢裂殖酵母)与产丁酸细菌之间的协同交叉喂养网络,是预测患者对免疫治疗响应率和生存期的强有力生物标志物。
然而,科学的严谨在于绝不回避风险。论文深刻指出,活体细菌进入人体后绝非静止不变的化学分子,它们是受进化法则支配的动态生命体,“宿主内进化”是一把悬在活体疗法头顶的双刃剑。一项震撼性研究表明,即便是公认安全的罗伊氏乳杆菌,在小鼠体内单一定植时,也会在极短时间内于调控基因上积累突变。这种快速进化改变了细菌的细胞壁结构,使其获得了更强的免疫逃逸能力和潜在的致病性[11]。此外,基因工程菌还面临着将耐药基因或改造质粒通过水平基因转移传递给肠道常驻病原体的巨大风险。
为了驾驭这股狂野的生物学力量,科学家们正在构建严密的生物安全防御体系。在临床前阶段,研究人员开发了快速整合细菌进化分析框架,通过人为施加选择压力,在一个月内即可评估候选菌株向高毒力方向突变的风险。在基因工程层面,合成生物学家正在设计精密的自杀开关和合成营养缺陷型回路。这些机制确保工程菌一旦离开肠道特定环境,或在完成预定疗程后停止特定营养底物供应时,便会触发程序性细胞死亡。这种从基因底层锁死的安全阀,是确保下一代益生菌不会对人类生态圈造成不可逆破坏的终极保障。
迈向个性化活体药物时代,大模型赋能与临床重塑
下一代益生菌的崛起,正在深刻改变我们对疾病干预的认知。2023年,美国FDA历史性地批准了首批活体生物药,这标志着微生态疗法正式跨越了灰色地带,堂堂正正地步入了受严格监管的现代制药殿堂[12]。在这个激动人心的进程中,大型语言模型和多模态人工智能的深度介入,将以前所未有的速度推动活体药物的临床转化。
在重塑药物发现网络方面,人工智能展现出了跨越传统生物学试错屏障的强大能力。基于图神经网络的预测模型通过深度整合Probio数据库与庞大的ICD-10疾病代码库,能够在错综复杂的生物学网络中精准预测老菌株的新用途。算法通过网络拓扑分析,成功预测了詹氏乳杆菌在治疗克罗恩病和溃疡性结肠炎中的潜在临床价值。这种计算机模拟预测极大地缩小了候选菌株的搜索空间,使得科学家能够绕过漫长且昂贵的盲目筛选,直接进入高价值的靶向验证阶段,从而极大地缩短了临床前的试错周期。
与此同时,制剂的稳定性与临床试验设计的复杂性一直是横亘在益生菌工业化道路上的两座大山。如今,主动机器学习正被创新性地用于攻克益生菌制剂不稳定的工业瓶颈。面对成百上千种潜在的药物辅料,传统方法只能依赖耗时耗力的物理混合测试。而主动机器学习模型能够通过极少量的初始湿实验反馈,迅速计算出不同辅料组合对特定菌株活性的保护概率,并在几轮迭代内锁定最佳的配方组合。在临床试验的顶层设计环节,大型语言模型同样扮演着不可或缺的科研副驾驶角色。研究表明,诸如ChatGPT-4o等先进的语言模型,能够以高达95%的准确率从海量文献中提取和优化复杂的临床试验方案,不仅能精准识别统计分析计划和药代动力学指南中的关键监管要求,还能协助研究人员评估最佳的采样窗口与纵向研究设计,从而大幅提升了临床转化的效率与成功率。
在个性化定植预测领域,人工智能同样大放异彩。利用群落规模代谢模型创建的个性化“入侵测定”框架,能够精确模拟个体肠道内的代谢盲区。例如,模型可以识别出艰难梭菌对琥珀酸或鸟氨酸的代谢依赖,从而精确计算出哪种下一代益生菌组合能最有效地与病原体竞争这些核心营养源,实现真正的量体裁衣。未来的医生在开具处方时,将不再仅仅依赖症状,而是基于患者的基因组、饮食习惯和基线肠道菌群数据,利用计算模型精确预测最佳的活体药物组合。
跨越现实鸿沟,普世价值与健康生活的新哲学
尽管前景广阔,我们仍需跨越现实的鸿沟。人工智能模型严重依赖于高质量的训练数据,且存在“幻觉”的固有风险;缺乏生物学机制约束的纯算法预测往往难以在湿实验中复现。此外,从动物模型到人类的转化依然充满不确定性,人类肠道极度个性化的定植抵抗机制使得疗效的普适性面临挑战。活体厌氧菌的规模化生产、冷链运输以及药代动力学的标准化评估,仍是横亘在产业化面前的巨大壁垒。
从更宏观的社会视角来看,下一代益生菌技术不仅是一场医疗技术的革新,更是一次人类健康哲学的升华。它教会我们以“共生”而非“对抗”的眼光看待生命。我们体内的微生物不再是需要被强力抗生素无差别清除的潜在威胁,而是维系生命稳态不可或缺的隐形器官。随着活体生物药技术的普及与人工智能算力的跃升,未来的处方上可能不再是一堆冰冷的化学分子式,而是为您量身定制的、充满活力的微型生物兵团。精准的微生态健康管理将融入我们的日常起居,从源头上构筑起抵御慢性疾病的坚固防线,让每一个人都能在微观世界的和谐共舞中,拥抱更加健康、充满活力的未来。
参考资料
[1]Suez, J., Zmora, N., Segal, E. & Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 25, 716–729 (2019).
[2]Caballero-Flores, G., Pickard, J. M. & Núñez, G. Microbiota-mediated colonization resistance: mechanisms and regulation. Nat. Rev. Microbiol. 21, 347–360 (2023).
[3]Suez, J. et al. Post-antibiotic gut mucosal microbiome reconstitution is impaired by probiotics and improved by autologous FMT. Cell 174, 1406–1423.e16 (2018).
[4]Heinken, A. et al. Genome-scale metabolic reconstruction of 7,302 human microorganisms for personalized medicine. Nat. Biotechnol. 41, 1320–1331 (2023).
[5]Silva-Andrade, C. et al. A machine-learning approach for predicting butyrate production by microbial consortia using metabolic network information. PeerJ 13, e19296 (2025).
[6]Cheng, A. G. et al. Design, construction, and in vivo augmentation of a complex gut microbiome. Cell 185, 3617–3636.e19 (2022).
[7]Depommier, C. et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat. Med. 25, 1096–1103 (2019).
[8]Li, M. et al. Engineered probiotics with sustained release of interleukin-2 for the treatment of inflammatory bowel disease after oral delivery. Biomaterials 309, 122584 (2024).
[9]Cao, F. et al. Engineering clinically relevant probiotics with switchable ‘nano-promoter’ and ‘nano-effector’ for precision tumor therapy. Adv. Mater. 36, e2304257 (2024).
[10]Sokol, H. et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl Acad. Sci. USA 105, 16731–16736 (2008).
[11]Yang, Y. et al. Within-host evolution of a gut pathobiont facilitates liver translocation. Nature 607, 563–570 (2022).
[12]Feuerstadt, P. et al. SER-109, an oral microbiome therapy for recurrent Clostridioides difficile infection. N. Engl. J. Med. 386, 220–229 (2022).+
文章由麦基洗德生物工程有限公司研发中心专家团队基于国际权威期刊新近发表的文献进行客观分析解读,旨在促进学术交流与行业洞察。文中引用的图表、数据均来自原始文献,版权归原作者及出版方所有。如涉及版权问题,请及时联系我们,我们将第一时间处理。
